что такое ВПР на семейном образовании, как подготовить к ним ребёнка
Что такое ВПР
Всероссийские проверочные работы (ВПР) — это контрольные работы с едиными стандартизированными заданиями, которые проверяют знания школьников по тому или иному предмету.
ВПР — это не аттестация. Результат этих проверочных работ не должен влиять на годовую оценку.
С помощью Всероссийских проверочных работ оценивают соответствие знаний учеников школы федеральным государственным образовательным стандартам (ФГОС). Результаты ВПР показывают, нужна ли образовательной организации помощь, которая может выражаться, например, в дополнительных курсах повышения квалификации для педагогов.
На каждый предмет отведён свой день. Одна контрольная занимает 45 минут. Тестовых заданий нет, участник ВПР должен сам вписать короткий или расширенный ответ. Даты проведения ВПР строго регламентированы.
Список предметов и классов каждый год меняется.
По каким предметам проводится ВПР?
Предметы в 4 классе:
- русский язык (диктант и тестирование),
- математика,
- окружающий мир.

В 4 классе ВПР — обязательный срез знаний для перехода в среднюю школу.
Предметы в 5 классе:
- русский язык,
- математика,
- история,
- биология.
Предметы в 6 классе:
- русский язык,
- математика,
- история,
- обществознание,
- биология,
- география.
Предметы в 7–8 классах:
- русский язык,
- математика,
- иностранный язык,
- история,
- обществознание,
- физика,
- химия,
- биология,
- география.
В 10 классе только один предмет для проведения всероссийских проверочных работ — география.
Предметы в 11 классе:
- химия,
- математика,
- иностранный язык,
- история,
- физика,
- биология.
Нужно ли сдавать ВПР хоумскулеру
Формально дети на семейном образовании не обязаны сдавать Всероссийские проверочные работы, ведь это контроль качества образования в школе, а «семейники» не находятся в её контингенте.
Однако образовательная организация может потребовать это от ребёнка на семейном образовании в момент сдачи аттестаций. В этот период хоумскулер прикреплён к школе и находится в статусе экстерна. Напомним, что экстернат — это форма аттестации на семейном образовании и самообразовании. В таком случае проверочные работы придётся писать, но можно договориться, чтобы результаты ВПР пошли в зачёт промежуточной аттестации.
Как подготовиться к ВПР
Ничего страшного в сдаче ВПР нет — это отличный срез знаний. С помощью проверочных работ можно определить сильные и слабые стороны хоумскулера и скорректировать образовательную траекторию.
Кроме того, Всероссийские проверочные работы помогают детям психологически привыкнуть к подобным испытаниям. Этот навык пригодится хоумскулеру на государственной итоговой аттестации — ОГЭ и ЕГЭ. Рекомендуется пройти ВПР в 8, 10 и 11 классах для тренировки.
Специально готовиться ребёнку к Всероссийским проверочным работам не нужно, ведь они рассчитаны на базовый уровень подготовки школьника. Достаточно хорошо заниматься весь год.
Но сам формат может быть непривычен ребёнку, который впервые участвует в ВПР. Поэтому стоит заранее посмотреть задания прошлых лет и попробовать их решить.
Получать комплексные знания по школьной программе можно в домашней онлайн-школе «Фоксфорда». У нас проходит обучение с 1 по 11 класс, а занятия ведут преподаватели из лучших вузов страны. В рамках индивидуальной программы обучения ребёнок может изучать любимые предметы на углублённом уровне. Кроме того, существуют специальные курсы, где можно опробовать формат Всероссийских проверочных работ.
ВПР 2020. Классы. Предметы
Всероссийские проверочные работы ВПР в 2020 году пройдут в 4, 5, 6, 7, 8, 10, 11 классах.
Участие 4, 5, 6, 7 классов в проверочной работе будет являться обязательным для всех школ.
В 8 классах ВПР 2020 пройдут в режиме апробации.
Для 10-11 классов участие в данной контрольной — добровольное.
ВПР 2020 год — классы и предметы
Источник информации:
— Официальный сайт Рособрнадзора
— Презентация с пресс конференции Рособрнадзора
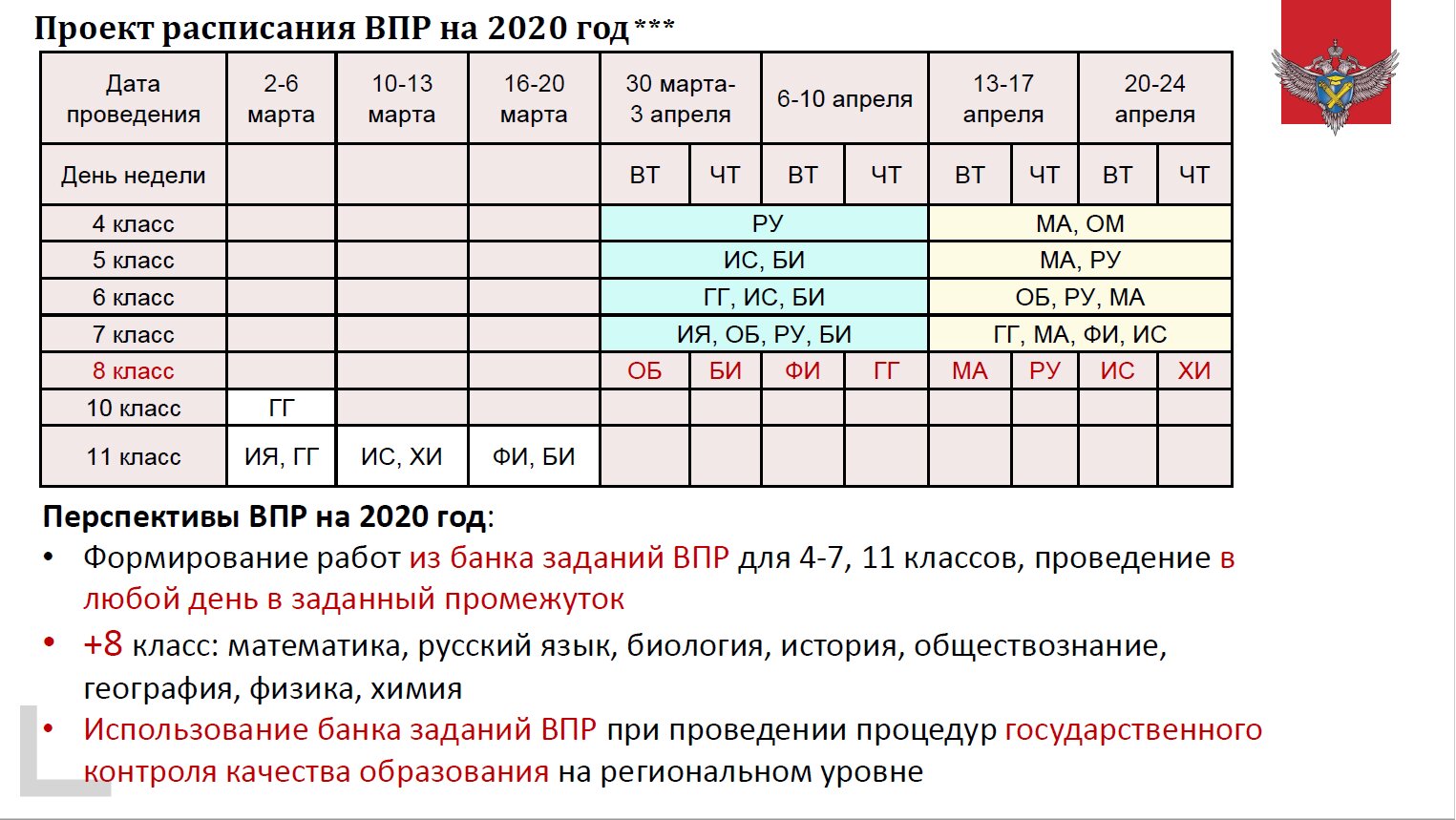
Проект расписания ВПР 2020
4 класс
— русский язык (часть 1- диктант, часть 2)
— математика
— окружающий мир
5 класс (ВПР 2020 обязательны для всех)
— русский язык
— математика
— история
— биология
6 класс (обязательны для всех)
— русский язык
— математика
— история
— биология
— география
— обществознание
7 класс (ВПР 2020 впервые пройдут в обязательном порядке)
— иностранный язык
— обществознание
— русский язык
— биология
— математика
— география
— история
— физика
8 класс ( в режиме апробации)
математика,
русский язык,
биология,
история,
обществознание,
география,
физика,
химия
10 класс
— география
11 класс
— история
— биология
— география
— химия
— физика
— иностранный язык
Нововведения ВПР на 2020 год:
— Формирование работ из банка заданий ВПР для 4-7, 11 классов, проведение в любой день в заданный промежуток
— Использование банка заданий ВПР 2020 при проведении процедур государственного контроля качества образования на региональном уровне.
Смотрите также:
ВПР-2021
Расписание проведения ВПР-2021 в 8 классах лицея:
- 02.04.2021 г. — предмет № 1 (предмет определяется федеральным координатором) —8 А — физика, 8 Б — биология, 8 В — химия.
- 06.04.2021 г. — русский язык — 8 А, Б, В классы.
- 08.04.2021 г. — предмет № 2 (предмет определяется федеральным координатором) —
- 14.04.2021 г. — математика — 8 А, Б, В классы.
Федеральная служба по надзору в сфере образования и науки утвердила расписание проведения всероссийских проверочных работ (ВПР) в 2021 году.
Согласно данному документу, ВПР пройдут для обучающихся 4-8 классов в штатном режиме, для обучающихся 11 классов – по решению школы. Конкретные даты проведения ВПР для каждого класса и предмета школы определят самостоятельно в рамках установленного расписанием периода.
Обучающиеся 8 классов с 15 марта по 21 мая напишут ВПР по русскому языку и математике, а также по двум предметам на основе случайного выбора. Восьмиклассникам могут встретиться ВПР по истории, биологии, географии, обществознанию, физике, химии. Информация о распределении предметов по классам в каждой параллели будет направлена школам через их личные кабинеты в Федеральной информационной системе оценки качества образования.
Информация о ВПР-2021 для учащихся
Информация о ВПР-2021 для родителей
Всероссийские проверочные работы (ВПР)
График проведения ВПР-2021
|
редмет (ВПР) |
4 класс |
5 класс |
6 класс |
7 класс |
8 класс |
11класс |
|
Русский язык |
13 апреля, 15 апреля |
15 апреля |
13 апреля | 8 апреля |
19 апреля |
— |
|
Математика |
20 апреля |
8 апреля |
19 апреля | 6 апреля | 13 апреля |
— |
|
Окружающий мир |
22 апреля |
— |
— |
— |
— |
— |
|
История |
— |
6 апреля |
15/21 апреля | 13 апреля | 8/15 апреля | 3 марта |
|
Биология |
— |
13 апреля |
15/21 апреля | 19 апреля | 8/15 апреля | 1 марта |
|
География |
— |
— |
15/21 апреля |
21 апреля | 8/15 апреля | 2 марта |
|
Обществознание |
— |
— |
15/21 апреля |
23 апреля | 8/15 апреля | 1 марта |
|
Английский язык |
— |
— |
— |
26-30 апреля |
— |
1 марта |
|
Химия |
— |
— |
|
— |
8/15 апреля |
2 марта |
|
Физика |
— |
— |
— |
15 апреля |
8/15 апреля | 2 марта |
В 6 и 8 классах указаны две даты 15/21 и 8/15 апреля соответственно , т.к. обучающиеся будут выполнять работы по двум предметам на основе случайного выбора.
Результаты Всероссийских проверочных работ 2020 года
|
Предмет |
Кол-во участников |
«5» |
«4» |
«3» |
«2» |
Кач-во |
Рез-ть |
5-е классы (127 человек) |
|||||||
|
Русский язык |
111 |
31 |
53 |
24 |
3 |
76% |
97% |
|
Математика |
102 |
37 |
48 |
16 |
1 |
83% |
99% |
|
Окружающий мир |
105 |
26 |
69 |
10 |
0 |
90% |
100% |
|
6-е классы (150 человек) |
|||||||
|
Русский язык |
132 |
27 |
55 |
42 |
8 |
62% |
94% |
|
Математика |
123 |
41 |
49 |
30 |
3 |
73% |
97,5% |
|
История |
127 |
28 |
66 |
33 |
0 |
74% |
100% |
|
Биология |
130 |
15 |
61 |
50 |
4 |
58% |
97% |
|
7-е классы (132 человека) |
|||||||
|
Русский язык |
96 |
3 |
24 |
40 |
29 |
28% |
70% |
|
Математика |
114 |
31 |
53 |
22 |
8 |
74% |
93% |
|
История |
97 |
5 |
60 |
32 |
0 |
67% |
100% |
|
Биология |
113 |
2 |
58 |
46 |
7 |
53% |
94% |
|
География |
110 |
26 |
64 |
19 |
1 |
82% |
99% |
|
Обществознание |
117 |
12 |
35 |
65 |
5 |
41% |
96% |
|
8-е классы (100 человек) |
|||||||
|
Русский язык |
85 |
14 |
46 |
20 |
5 |
71% |
94% |
|
Математика |
79 |
33 |
29 |
16 |
1 |
78% |
99% |
|
История |
85 |
9 |
58 |
18 |
0 |
79% |
100% |
|
Биология |
81 |
1 |
43 |
35 |
2 |
54% |
97,5% |
|
География |
82 |
9 |
38 |
35 |
0 |
57% |
100% |
|
Обществознание |
82 |
24 |
43 |
12 |
3 |
82% |
96% |
|
Физика |
85 |
31 |
39 |
15 |
0 |
82% |
100% |
|
Английский язык |
86 |
25 |
37 |
23 |
1 |
72% |
99% |
|
9-е классы (119 человек) |
|||||||
|
Русский язык |
97 |
13 |
50 |
24 |
10 |
65% |
90% |
|
Математика |
|||||||
График проведения ВПР-2020
График проведения ВПР 2020
|
Предмет (ВПР) |
4 класс |
5 класс |
6 класс |
7 класс |
8 класс |
11класс |
|
Русский язык |
7 апреля, 9 апреля |
14 апреля |
13 апреля |
9 апреля |
16 апреля |
— |
|
Математика |
14 апреля |
16апреля |
20 апреля |
22 апреля |
14 апреля |
— |
|
Окружающий мир |
16 апреля |
— |
— |
— |
— |
— |
|
История |
— |
7 апреля |
9 апреля |
17 апреля |
21 апреля |
10 марта |
|
Биология |
— |
9 апреля |
10 апреля |
8 апреля |
2 апреля |
18 марта |
|
География |
— |
— |
7 апреля |
15 апреля |
9 апреля |
— |
|
Обществознание |
— |
— |
15 апреля |
7 апреля |
31 марта |
— |
|
Английский язык |
— |
— |
— |
10 апреля |
— |
2 марта |
|
Химия |
— |
— |
— |
— |
23 апреля |
12 марта |
|
Физика |
— |
— |
— |
20 апреля |
7 апреля |
16 марта |
Результаты Всероссийских проверочных работ 2018-2019 учебного года
|
Предмет |
«5» |
«4» |
«3» |
«2» |
%успеваемости |
%качества |
|
4 класс |
||||||
|
Математика |
64 |
71 |
4 |
— |
100% |
97% |
|
Русский язык |
35 |
73 |
32 |
— |
100% |
77% |
|
Окружающий мир |
59 |
71 |
6 |
— |
100% |
96% |
|
5 класс |
||||||
|
Биология |
1 |
74 |
38 |
— |
100% |
66% |
|
История |
32 |
53 |
37 |
8 |
94% |
65% |
|
Математика |
58 |
40 |
9 |
— |
100% |
92% |
|
Русский язык |
26 |
37 |
47 |
21 |
84% |
48% |
|
6 класс |
||||||
|
Биология |
2 |
72 |
24 |
1 |
99% |
75% |
|
География |
16 |
70 |
10 |
— |
100% |
90% |
|
История |
30 |
53 |
10 |
2 |
99% |
87% |
|
Математика |
18 |
60 |
17 |
1 |
99% |
81% |
|
Обществознание |
23 |
64 |
10 |
1 |
99% |
89% |
|
Русский язык |
22 |
43 |
30 |
4 |
96% |
66% |
|
7 класс |
||||||
|
Биология |
15 |
73 |
28 |
3 |
97% |
74% |
|
География |
11 |
63 |
38 |
— |
100% |
66% |
|
История |
25 |
62 |
21 |
2 |
98% |
79% |
|
Математика |
52 |
53 |
10 |
— |
100% |
91% |
|
Обществознание |
3 |
68 |
41 |
2 |
98% |
62% |
|
Русский язык |
18 |
58 |
32 |
12 |
96% |
63% |
|
Физика |
1 |
50 |
57 |
6 |
95% |
45% |
Расписание ВПР-2019
| Предмет (ВПР) | 4 класс | 5 класс | 6 класс | 7 класс | 11 класс |
| Русский язык | 15 апреля- 19 апреля 2019 |
25 апреля 2019 |
23 апреля 2019 | 9 апреля 2019 | |
| Математика | 22-26 апреля 2019 | 23 апреля 2019 | 25 апреля 2019 | 18 апреля 2019 | — |
| Окружающий мир | 22-26 апреля 2019 | — | — | — | |
| История | — | 16 апреля 2019 | 11апреля 2019 | 25 апреля 2019 | 13 марта |
| Биология | — | 18 апреля 2019 | 16 апреля 2019 | 11 апреля 2019 | 21 марта |
| География | — | — | 9 апреля 2019 | 16 апреля 2019 | 12 марта |
| Обществознание | — | — | 18 апреля 2019 | 4 апреля 2019 | — |
| Английский язык | — | — | — | 2 апреля 2019 | 5 марта |
| Химия | — | — | — | 14 марта | |
| Физика | — | — | — | 19 марта |
У учеников 9 классов по учебному предмету «русский язык» в форме ВПР -итоговое собеседование с 14 по 16 февраля 2019 года.
Страница 1 из 2
Всероссийские проверочные работы
https://fioco.ru/ru/osoko/vpr/ — Федеральный институт оценки качества образования (поддержка ВПР)
Документы:
НОВОЕ!!!
График проведения ВПР в 2021 году
ГБОУ гимназия № 498
Невского района Санкт-Петербурга
Класс | Предмет | Дата | ||
Март | Апрель | Май | ||
4 класс | Русский язык I |
| 08.04. |
|
Русский язык II |
| 13.04 |
| |
Математика |
| 20.04 |
| |
Окруж.мир |
| 22.04 |
| |
5 класс | Русский язык |
| 13.04 |
|
Математика |
| 22.04 |
| |
История |
|
| 14.05 | |
Биология |
| 20.04 |
| |
| ||||
6 класс | Русский язык |
| 06.04 |
|
Математика |
| 16.04 |
| |
Предмет по выбору |
| 27.04 |
| |
Предмет по выбору |
|
| 13.05 | |
| ||||
7 класс | Русский язык |
| 14.04 |
|
Математика |
| 23.04 |
| |
История |
| 28.04 |
| |
Биология |
|
| 11.05 | |
География |
| 01.04 |
| |
Обществознание |
| 07.04 |
| |
Физика |
| 29.04 |
| |
Англ.яз. |
| 15.04 |
| |
| ||||
8 класс | Русский язык |
| 02.04 |
|
| Математика |
| 08.04 |
|
| Предмет по выбору |
| 21.04 |
|
| Предмет по выбору |
|
| 12.05 |
| ||||
10 класс | География | 10.03 |
|
|
| ||||
11 класс | История | 11.03 |
|
|
| Биология | 12.03 |
|
|
| География | 17.03 |
|
|
| Физика | 18.03 |
|
|
| Химия | 10.03 |
|
|
| Английский язык | 16.03 |
|
|
| ||||
Распоряжение о НИКО. Скачать
Письмо Рособрнадзора от 05.08.2020 №13-404 «О проведении всероссийских проверочных работ в 5-9 классах осенью 2020 года»
Информационное письмо заместителя руководителя Федеральной службы по надзору в сфере образования и науки (Рособрнадзор) от 04.09.2020 № 13-444 «Об организации проведения всероссийских проверочных работ в 5-9 классах осенью 2020 года» Скачать
Письмо Рособрнадзора от 16.11.2016 №05-628 «О проведении Всероссийских проверочных работ в 11 классе». Скачать
Письмо Рособрнадзора от 13.12.2016 №05-714 «В дополнение к письму Рособрнадзора от 16.11.2016 №05-628». Скачать
Письмо Рособрнадзора от 02.02.2017 № 05-41 «О Всероссийских проверочных работах». Скачать
Проведение Всероссийских проверочных работ (далее — ВПР) организовано с целью формирования единого образовательного пространства в Российской Федерации. Варианты работ и система оценивания разрабатываются на федеральном уровне и дают возможность оценить учебные результаты обучающихся по единым критериям.
Обращаем внимание, что результаты ВПР не должны учитываться при выставлении годовых отметок по предметам или при получении аттестата о среднем общем образовании.
Согласно Распоряжению Рособрнадзора от 06.12.2016 года № 3167-05 ВПР по предмету география является обязательным для всех учащихся.
Федеральный институт оценки качества образования (ФИОКО) опубликует на своем сайте образцы и описания всероссийских проверочных работ (ВПР), которые пройдут в 2020 году.
Образцы и описания проверочных работ дают возможность учащимся и преподавателям составить представление о структуре ВПР, количестве заданий, их форме и уровне сложности, перечне вопросов и тем, которые могут встретиться в ВПР, системе оценивания.ВПР представляют собой контрольные работы, которые проводятся в общеобразовательных организациях по завершении обучения в каждом классе. Это итоговые контрольные работы, которые проходят по отдельным учебным предметам для оценки уровня подготовки школьников с учетом требований федеральных государственных образовательных стандартов. Их организация предусматривает единое расписание, использование единых типов заданий и единых критериев оценивания.
Учащиеся четвертых классов напишут ВПР по русскому языку, математике и окружающему миру, пятиклассники – по русскому языку, математике, биологии и истории, шестиклассники – по русскому языку, математике, биологии, истории, обществознанию и географии, семиклассники – по русскому языку, математике, биологии, истории, обществознания, географии и английскому языку, одиннадцатиклассники – по географии, физике, химии, биологии, истории и иностранному языку.
ФИПИ публикует описания и образцы вариантов для проведения в 11 классах Всероссийских проверочных работ (ВПР) по учебным предметам:
БИОЛОГИЯ-11 (скачать)
ГЕОГРАФИЯ-11 (скачать)
ИСТОРИЯ-11 (скачать)
ФИЗИКА-11 (скачать)
ХИМИЯ-11 (скачать)
По каждому из предметов опубликованы:
— описание ВПР по данному предмету, включающее Кодификатор элементов содержания и требований к уровню подготовки выпускников образовательных организаций для проведения ВПР. Кодификаторы составлены на основе Федерального компонента государственных стандартов основного общего и среднего (полного) общего образования для изучения предмета на базовом уровне. В описании ВПР содержатся сведения о распределении заданий в работе по блокам содержания и проверяемым способам действий, о системе оценивания отдельных заданий и работы в целом.
— образец всероссийской проверочной работы, который дает представление о структуре всероссийской проверочной работы, количестве и форме заданий, уровне их сложности.
— ответы и критерии оценивания к образцу проверочной работы.
Что такое ВПР (Всероссийская проверочная работа)?
Статьи
Линия УМК Мерзляка. Математика. Алгебра. Геометрия (5-11) (Б)
Линия УМК География. «Классическая линия» (5-9)
Линия УМК В. П. Дронова. География (Роза ветров) (10-11) (Б)
Линия УМК Волобуева-Пономарева. Всеобщая история (10-11) (БУ)
Биология
Зачем создана, когда проводится, чем грозит, как подготовиться к ней психологически? Обращаемся к официальным данным и интервью представителей Рособрнадзора.
25 июля 2017
Всероссийские проверочные работы (ВПР) — практика, призванная наладить регулярную проверку уровня знаний школьников на соответствие федеральным государственным образовательным стандартам. Первые ВПР российские школьники написали в 2015 году. С 2016 года проведение Всероссийских проверочных работ стало регулярным.
Изучив официальные источники, мы отвечаем на вопросы о ВПР, которые чаще всего задают родители.
Основные сведения: зачем, для кого, как
Зачем потребовалось вводить ВПР? Неужели ЕГЭ не дает всех необходимых сведений?
Профильные ЕГЭ по всем предметам учащиеся сдают не в общем порядке, а по выбору, то есть, нет никакой возможности получить полные данные о том, как усвоили предмет остальные учащиеся. К тому же выпускники осознают, что от результатов ЕГЭ зависит их будущая учеба, карьера, поэтому очень тщательно готовятся к профильным экзаменам, чаще всего прибегая к помощи репетиторов и посещая спецкурсы. Поэтому по результатам ЕГЭ нельзя судить о том, какова роль школы в успехе выпускников. Для того чтобы получить более прозрачную картину, введены Всероссийские проверочные работы, составленные по единым требованиям.
В 11 классах ВПР проводятся только для тех учеников, которые не выбрали ЕГЭ по соответствующим предметам. То есть кто сдает ЕГЭ по географии, тот не пишет ВПР по географии.
«Сейчас школы сами проверяют, как учащиеся освоили предметы, не входящие в ЕГЭ или ГИА-9, сами проводят контрольные работы. Проводят они их по собственным заданиям. Никто не контролирует, как эти задания составлены, каков их уровень сложности, как оценивается их выполнение. Вводя ВПР, мы предложили школам единый стандарт оценки…» — Анзор Музаев, заместитель руководителя Федеральной службы по надзору в сфере образования и науки, в интервью МИА «Россия сегодня» от 14.12.2016.
Чтобы продолжить чтение, авторизуйтесь на сайте.
МОУ «СОШ №2 г. Ртищево Саратовской области»
2020 -2021 учебный год
Результаты исследования объективности системы оценивания образовательных результатов в образовательных организациях Саратовской области в 2020/2021 учебном году
Выписка из протокола №6 от 26.03.21г. совещания при директоре
ГРАФИК ПРОВЕДЕНИЯ ВПР -2021
№ | Предмет | Класс | Дата проведения |
| Обществознание | 7 | 16 марта |
| Русский язык (часть 1) | 4 | 16 марта |
| Предмет 1 | 6 | 18 марта |
| Русский язык (часть 2) | 4 | 18 марта |
| Предмет 1 | 8 | 19 марта |
| История | 5 | 22 марта |
| Физика | 7 | 22 марта |
| Предмет 2 | 6 | 23 марта |
| Предмет 2 | 8 | 1 апреля |
| История | 7 | 2 апреля |
| Биология | 5 | 2 апреля |
| Русский язык | 8 | 5 апреля |
| Математика | 6 | 5 апреля |
| Английский язык | 7 | 6 апреля |
| Математика | 4 | 6 апреля |
| Русский язык | 5 | 7 апреля |
| Английский язык | 7 | 7 апреля |
| Биология | 7 | 8 апреля |
| Математика | 8 | 9 апреля |
| Русский язык | 7 | 13 апреля |
| Математика | 5 | 13 апреля |
| Окружающий мир | 4 | 14 апреля |
| Математика | 7 | 15 апреля |
| Русский язык | 6 | 15 апреля |
| География | 7 | 16 апреля |
Образцы и описания проверочных работ для проведения ВПР в 2021 году
Расписание ВПР 2021
НОВОСТИ ВПР 2021
План- график ВПР с изменениями
Порядок проведения ВПР
Время проведения ВПР
Методические рекомендации на основе результатов ВПР , проведенных в сентябре — октябре 2020г. утвержденные МИНПРОСВЕЩЕНИЕМ РОССИИ
План мероприятий («дорожная карта») по реализации образовательных программ начального общего и основного общего образования в МОУ «СОШ № 2 г. Ртищево Саратовской области» на основе результатов ВПР, проведенных в сентябре-октябре 2020 г.
_________________________________________________________________________________________________________________________________________
2019-2020 учебный год
Проведение всероссийских проверочных работ (ВПР) для учащихся 4-8 классов
перенесено на осень 2020 года, планируется, что они пройдут в сентябре-октябре (ссылка).
Проведение Всероссийских проверочных работ будет перенесено с конца этого учебного года на осень. Об этом Министр просвещения Сергей Кравцов заявил сегодня в ходе брифинга, посвящённого изменениям в вопросах оценки качества общего образования в 2019/2020 учебном году.
Решение о переносе сроков проведения ВПР было принято по итогам консультаций с педагогами, регионами, представителями образовательного сообщества и родительским сообществом.
Министр просвещения подчеркнул, что ВПР – это не формальная аттестация самих школьников: их результаты нужны для корректировки образовательного процесса. Оценки за ВПР выставляться не будут.
«Такая диагностика должна показать уровень знаний школьников и выявить возможные пробелы в знаниях. После проведения этих диагностических процедур школы, учителя получат соответствующие рекомендации, и будет выстроена необходимая методическая работа. К этой работе мы привлечём педагогические вузы, институты повышения квалификации, методические службы. Она будет координироваться Министерством просвещения», – рассказал Министр.
Особенности организации проведения ВПР-2020 по иностранному языку
Проект расписания ВПР 2020
Подготовка к ВПР -2020
Демоверсии → 4 класс | 5 класс | 6 класс | 7 класс | 8 класс
Демоверсии 11 класс
2018-2019 учебный год
Ирина Седова рассказала о новшествах проведения Всероссийских проверочных работ в 2018/2019 году
График проведения ВПР в 2018/2019 году (ПРОЕКТ)
Приказ от 29.01.2019 №84 «О проведении Федеральной службой по надзору в сфере образования и науки мониторинга качества подготовки обучающихся общеобразовательных организаций в 2019 г.»
Приказ №104 от 07.02.2019г. «О внесении изменений в график проведения Федеральной службой по надзору в сфере образования и науки мониторинга качества подготовки обучающихся»
План график школы по подготовке к проведению ВПР
Демоверсии ВПР-2019 для 4 класса
Демоверсии ВПР-2019 для 5 класса
Демоверсии ВПР-2019 для 6 класса
Демоверсии ВПР-2019 для 7 класса
Демоверсия ВПР -2019 для 10 класса
_______________________________________________________________________________________________________________________________________________________________________________________
2017-2018 учебный год
Письмо и приказ о проведении ВПР
План-график
Порядок проведения
Всероссийские проверочные работы
ФГБНУ «ФИПИ» публикует описания и образцы вариантов для проведения в 11 классах Всероссийских проверочных работ (ВПР) 2018 года по 6 учебным предметам: истории, географии, биологии, физике, химии и иностранным языкам (английский, немецкий и французский языки).
Демоверсии ВПР 2018 для 4 класса ВПР Официальные образцы заданий и описание работ.
Ссылка: http://4ege.ru/vpr/55620-demoversii-vpr-2018-dlya-4-klassa.html
Демоверсии ВПР 2018 для 5 класса ВПР Новые демоверсии 2018 года для пятиклассников.
ссылка: http://4ege.ru/vpr/55621-demoversii-vpr-2018-dlya-5-klassa.html
Демоверсии ВПР 2018 по всем предметам для 6 класса
ВПР Официальные демоверсии Всероссийской проверочной работы для 6 класса от СтатГрад.
ссылка: http://4ege.ru/vpr/55883-demoversii-vpr-2018-po-vsem-predmetam-dlya-6-klassa.html
Приказ №873 от 05.09.2017г.
ВПР 2 кл. образец
ВПР 5 кл. образец
Приказ по ВПР УОО №72 от 09.02.2017г.
Приказ ВПР по МОУ СОШ№2
Объявление
VPR — напоминание о регистрации личных исследований на людях в интеграторе EHS
Дата: 12 февраля 2021 г.
Attn: Йельские исследователи исследований на людях и доверенных лиц
Резюме: напоминание о регистрации Личные исследования на людях на интеграторе EHS
Уважаемые коллеги,
Поскольку количество случаев COVID продолжает оставаться высоким, мы пишем, чтобы напомнить вам, что все исследования, требующие личного взаимодействия с участниками исследования, будь то в университетском городке или за его пределами, включая участников исследования дома, требуют одобрения Человека-субъекта. Исследовательский комитет (HSRC).Это включает в себя активацию новых исследований, повторную активацию исследований, которые были приостановлены, исследования, переходящие от удаленных к личным визитам, и исследования, которые продолжались во время пандемии с личным взаимодействием.
Если вы еще не получили одобрение HSRC, вы должны как можно скорее отправить свои планы безопасности через EHS Integrator (требуется VPN). В это время высокой передачи особенно важно зарегистрировать свое исследование в EHS Integrator, чтобы вы могли получать важные обновления и рекомендации по безопасности при проведении личных исследований на людях.
Обратите внимание, что проверка HSRC не является частью процесса проверки IRB и вспомогательного комитета. Следовательно, одобрение HSRC не следует толковать как одобрение IRB на проведение исследований, которые еще не получили одобрения IRB. Кроме того, одного одобрения IRB недостаточно для проведения исследования при личном общении во время реактивации исследования Йельского университета.
Спасибо за ваше постоянное сотрудничество и бдительность в эти трудные времена. При вашей поддержке и сотрудничестве мы сможем обеспечить максимально безопасную среду как для исследователей, так и для участников.
С уважением,
Майкл К. Крейр, доктор философии
Уильям Зиглер III Профессор нейробиологии
Заместитель проректора по исследованиям
Линда Коулман, доктор медицинских наук
Директор Программы защиты человеческих исследований
Офис управления исследованиями
Эрик Веласкес, доктор медицины
Роберт В. Берлинер, профессор медицины
Сопредседатель Комитета по исследованиям на людях
Грегори Маккарти, доктор философии.
Генри Форд II Профессор психологии
Сопредседатель Комитета по исследованиям на людях
COVID и исследования на людях
6 января 2021 г.
Отправлено от имени Марка А.Барто, вице-президент по исследованиям
За последние семь месяцев мы смогли возобновить множество исследовательских проектов, в том числе с участием людей. Каждый из них был рассмотрен и одобрен на кафедре, колледже и, в случае исследования с участием человека, на уровне университета с учетом рисков, преимуществ, контекста и ситуации с COVID-19 для соответствующего местоположения. Представленные вами планы управления и инфекционного контроля продемонстрировали твердую приверженность безопасности исследователей, участников и наших сообществ.Вместе исследовательское сообщество Texas A&M добилось успеха в предотвращении распространения вируса с помощью исследовательской деятельности. Однако очень важно, чтобы мы не ослабляли бдительность и не ослабляли соблюдение протоколов безопасности COVID.
Сегодня мы оказались в иных обстоятельствах, чем месяц назад. Хотя распространение вакцин обнадеживает, скорее всего, пройдут месяцы, прежде чем большая часть населения университетского городка сможет быть вакцинирована. В то же время мы наблюдаем рекордные уровни положительных результатов на местном и государственном уровне, а также загруженности отделений интенсивной терапии.Тестирование по-прежнему имеет решающее значение, и я поздравляю наших коллег, работающих в Глобальном исследовательском комплексе здравоохранения, за то, что тестирование на основе слюны является жизненно важным компонентом наших возможностей тестирования.
В настоящее время мы не рассматриваем дополнительные ограничения или требования для исследований, в том числе с участием людей. Мы продолжим следить за ситуацией как в сообществе, так и среди наших сотрудников и студентов, проживающих в кампусе, которым необходимо пройти тестирование в течение следующих нескольких недель.Тем не менее, я хочу самым решительным образом призвать вас перейти от исследований с участием людей к виртуальным взаимодействиям и сократить или исключить личные взаимодействия, если это вообще возможно.
Если уровень заражения COVID продолжит расти среди населения, включая студентов и членов сообщества, из которых мы привлекаем участников, может возникнуть необходимость прекратить личные исследования с участием людей. Ряд учреждений уже сделали это. Переход к чисто виртуальным действиям сейчас может доставлять неудобства, но может помочь избежать отключения в будущем.
Благодарим вас за вашу приверженность продвижению знаний посредством исследований, причем безопасным и ответственным образом. Стандарты, которые вы поддерживаете, и поведение, которое вы моделируете, могут стать самым важным уроком для молодых умов, которые мы формируем.
Добровольная экспертная оценка (VPR) пересмотра и реализации НСПДСБ
На своих девятом и одиннадцатом совещаниях Конференция Сторон поручила Исполнительному секретарю содействовать, в том числе , процессу добровольного обзора осуществления НСПДСБ.На своем двенадцатом совещании Конференция Сторон в пункте 3 своего решения XII / 29 приняла к сведению документы UNEP / CBD / COP / 12/25 / Add.3 и UNEP / CBD / COP / 12 / INF / 24. подготовлен Исполнительным секретарем по предлагаемому экспериментальному процессу добровольного коллегиального обзора для НСПДСБ и поручил Исполнительному секретарю, при условии наличия ресурсов, разработать методологию для процесса добровольного коллегиального обзора и представить отчет Вспомогательному органу по осуществлению (UNEP / CBD / SBI / 1/10 / Add.1). На своем тринадцатом совещании Конференция Сторон в решении XIII / 25 приняла к сведению прогресс, достигнутый в разработке механизма добровольного коллегиального обзора, особенно в разработке проекта методологии обзора, и поручила Исполнительному секретарю , при наличии ресурсов, для облегчения дальнейшего тестирования и разработки методологии, в том числе ее применения на экспериментальном этапе, и для представления отчета о ходе работы, включая информацию о стоимости экспериментального этапа, Вспомогательному органу по внедрению. на своем втором заседании.На своем четырнадцатом совещании Конференция Сторон в решении 14/29 приветствовала прогресс, достигнутый в разработке механизма добровольного коллегиального обзора, а также положительный результат экспериментального этапа, инициированного решением XIII / 25, и постановила: включить добровольный экспертный обзор как элемент подхода к многомерному обзору в соответствии с Конвенцией. Правительства Японии, Норвегии и Швейцарии выделили средства и ресурсы на дальнейшее развитие процесса добровольной экспертной оценки.Была создана неофициальная рабочая группа для продолжения разработки и тестирования предлагаемой методологии (в общей сложности 17 Сторон назначили экспертов). Вводное совещание Рабочей группы было проведено в Тбилиси, Грузия, с 11 по 13 февраля 2015 года. На совещании обсуждалась методология, представленная двенадцатому совещанию Конференции Сторон, и были разработаны пересмотренная методология и план работы. Было также решено, что пересмотренная методология должна быть опробована как минимум в двух странах. Эфиопия и Индия были выбраны в качестве стран для тестирования, что дает хороший географический и социально-экономический диапазон (кроме того, обе страны недавно пересмотрели свои НСПДСБ).Второе совещание неофициальной рабочей группы было созвано в Берне, Швейцария, с 16 по 18 марта 2016 года для рассмотрения двух опытов тестирования и дальнейшей разработки методологии.
Общая цель процесса добровольного экспертного обзора Конвенции о биологическом разнообразии (КБР) — помочь Сторонам улучшить их индивидуальный и коллективный потенциал для более эффективного осуществления Конвенции. Конкретные цели заключаются в следующем:(a) Оценить разработку и осуществление национальных стратегий и планов действий по сохранению биоразнообразия (НСПДСБ) в контексте Стратегического плана в области сохранения и устойчивого использования биоразнообразия на 2011-2020 годы и подготовить конкретные рекомендации для рассматриваемых Сторон;
(b) предоставить возможности для взаимного обучения непосредственно вовлеченным Сторонам и другим Сторонам;
(c) Обеспечить большую прозрачность и подотчетность при разработке и реализации НСПДСБ перед общественностью и другими Сторонами.
Отчеты, подготовленные группами экспертов Эфиопии и Индии о процессе добровольной экспертной оценки, доступны ниже:VPR Обновление COVID-19 — 27 мая 2021 г. — вице-президент по исследованиям
8 июня 2021 г.
Уважаемые коллеги,
Спасибо всем за посещение Ратуши виртуальных исследований 24 мая, , . Мы надеемся, что ратуши VPR были информативными в общении с исследовательским кампусом. обновления, а также полезны для связи друг с другом во время этих сложных раз.Мы искренне ценим ваши отзывы и участие в этих мероприятиях мэрии.
Вот обновления VPR COVID-19:
- Запись Virtual Research Town Hall теперь доступна на U Box
- Обновлено руководство по маскам для лица в Университете здравоохранения штата Юта
U of U Health обновило руководство по использованию масок для лица в Университете здравоохранения штата Юта — в том числе в специализированных, многофункциональных и неклинических учреждениях (см. Приложение).Вы можете получить доступ к самой последней версии на Pulse. - COVID-19 Руководство по поездкам в университет
Напоминаем, что в настоящее время Университет ограничивает все входящие и исходящие бизнесы. до 30 июня 2021 г. Начиная с 1 июля ограничения на поездки внутри страны будут конец. Международные поездки подлежат процедурам утверждения. Посетите веб-сайт финансовых и бизнес-услуг для получения более подробной информации.
Еще раз благодарим вас за терпение, пока мы продолжаем просматривать эти обновления. и изменения. Пожалуйста, дайте нам знать, если у вас есть какие-либо вопросы. Мы с нетерпением ждем встречи все вы скоро.
С уважением,
Энди Вейрих, вице-президент по исследованиям
Дайан Патаки, заместитель вице-президента по исследованиям
Эрин Ротвелл, заместитель вице-президента по вопросам целостности исследований и соблюдения нормативных требований
изменений в С-концевой области дополнительного гена ВИЧ-1 vpr не дают клинических преимуществ субъектам, получающим нуклеозидную антиретровирусную терапию | Журнал инфекционных болезней
Аннотация
С-конец дополнительного белка вируса иммунодефицита человека типа 1 (ВИЧ-1) vpr действует в остановке вирусного клеточного цикла, ядерной локализации и апоптоза.Полиморфизмы в этой области описаны в ряде случаев длительного отсутствия прогрессирования. Мы определили vpr последовательностей заархивированных исходных образцов от 96 участников исторического испытания ингибиторов обратной транскриптазы с одним и двумя нуклеозидами. Затем эти последовательности анализировали по характеристикам начала исследования и результатов, таким как исходное абсолютное количество клеток CD4 + , предшествующее лечение, ответ клеток CD4 + и клинические конечные точки. Частота C-концевых мутаций не коррелировала с какими-либо показателями интенсивности заболевания.Изменения в этой части vpr не ослабляли болезнь.
Среди потенциальных вирусных детерминант клинического заболевания ВИЧ-1, дивергенция последовательностей в дополнительных белковых областях, а также в env и gag может влиять на прогрессирование заболевания [1–3]; например, изменения в HIV-1 nef предрасполагают к долгосрочному непрогрессированию (LTNP) [4, 5]. Отклонения от согласованной последовательности в других генах вспомогательного белка ВИЧ-1 были предложены в качестве дополнительных факторов, ослабляющих болезнь [1, 6].Vpr, один из 6 дополнительных белков ВИЧ, достаточно консервативен in vivo по сравнению с большинством лентивирусных белков. Хотя его роль в инфицировании ВИЧ-1 остается несколько неясной, in vitro он выполняет множество функций. Vpr останавливает клеточный рост в фазе G2 клеточного цикла, тем самым усиливая трансактивацию с помощью длинно-концевого повтора ВИЧ, и содержит сигналы ядерного импорта, которые способствуют захвату преинтеграционного комплекса ВИЧ [7]. Vpr также способствует нарушению трансмембранного потенциала митохондрий и активации цитохром с-зависимой каспазы [7–9].
Некоторые функции vpr, а именно, его апоптоз, ядерная локализация и способность останавливать клетки, картируются на его С-конце [8-11]. Мутационный анализ выявил область, охватывающую аминокислотные остатки 71–96, которые охватывают 2 жестко консервативных полипептидных мотива HFRIG, охватывающих обратный поворот β [10]. Недавнее сообщение продемонстрировало, что при применении к культурам лимфоцитарных клеток, мутант пептида vpr ВИЧ-1, псевдотипированного вирусом везикулярного стоматита по кодону 77, индуцирует меньший апоптоз, чем vpr дикого типа [9].Вирусные последовательности LTNPs содержат мутационные изменения в этой относительно консервативной области генома ВИЧ-1 [9, 12-14]; например, четыре пятых последовательностей vpr из 10 LTNP демонстрируют полиморфизм в кодоне 77 vpr , по сравнению с гораздо меньшим процентом в последовательностях из 15 не-LTNP [9]. Соответственно, мы исследовали, соответствует ли отклонение от консенсусных последовательностей C-конца клады B vpr ослабленному прогрессированию заболевания у участников клинических испытаний ВИЧ-1, получавших лечение до появления мощной антиретровирусной терапии.
Предметы, материалы и методы.
Субъекты, материалы и методы . Сто три ВИЧ-1-инфицированных субъекта без СПИДа, включенных в наш центр в течение 1991–92 гг. В большом многоцентровом слепом исследовании однократного обратного двойного нуклеозидного ингибитора обратной транскриптазы ( НИОТ) антиретровирусная терапия (протокол 175 Группы клинических исследований СПИДа) [15]. Информированное согласие было получено от пациентов, и были соблюдены руководящие принципы экспериментов на людях Министерства здравоохранения и социальных служб США.Для включения в исследование национальное родительское исследование требовало, чтобы субъект имел нормальный функциональный статус и исходное абсолютное количество Т-лимфоцитов CD4 + 200–500 клеток / мм 3 . Аликвоту базового уровня перед включением криоконсервированных мононуклеарных клеток периферической крови (PBMC), способных давать информацию о последовательности, получали от каждого из 96 субъектов в нашем центре. Данные об истории болезни, конечной точке и серийном абсолютном количестве CD4 + Т-лимфоцитов за 2 года были получены из базы данных исходного исследования [15].
Тотальную ДНК экстрагировали на спин-колонке (QIAamp DNA Mini kit; Qiagen) из криоконсервированных гранул первичных РВМС, а затем амплифицировали в объеме с помощью вложенной полимеразной цепной реакции (внешние праймеры NL4-3-552IF [5′-CCACCTTTGC-CTAGTGTTAG -3 ‘] и 5891R [5′-GCAGTTTTAGGCTGACT-TCC-3′], внутренние праймеры NL4-3-555 IF [5’-GAGG AC AG AT-GGAACAAGCC-3 ‘] и 5858R [5’-GGCTCTAGTCTAGGAT-CTAC- 3 ‘]) для области, кодирующей ВИЧ-1 vpr , кодоны 5–96.
Продукты очищали гель-электрофорезом и спин-колонкой (QIAquick gel kit; Qiagen), секвенировали дидезоксинуклеотидный цикл на ABI 377 и для анализа выравнивали с помощью программного обеспечения Sequencher с консенсусной последовательностью клады B.Клинические данные были получены после регистрации всех лабораторных результатов. Статистический анализ ассоциации по клиническим исходам полиморфизмов С-концевой последовательности vpr с использованием критерия Стьюдента t .
Результаты
Результаты. Современная эффективная комбинированная антиретровирусная терапия по стандарту лечения может маскировать преимущества, которые имеют ключевые мутации vpr на С-конце для клинического течения. По этой причине мы стремились ретроспективно оценить образцы и клинические данные из более раннего исследования.Протокол 175 ACTG был самой ранней доступной нам большой когортой с периода, предшествующего терапии тройными антиретровирусными комбинациями. Таким образом, он обладал преимуществом ограниченного воздействия лекарственного средства на исходном уровне. В этом многоцентровом исследовании 82% из почти 2500 участников были мужчинами, средний возраст которых составлял 35 лет. Среднее абсолютное количество CD4 + Т-клеток среди участников нашего сайта составило 331 клетка / мм 3 (диапазон: 186–533 клеток / мм 3 ; медиана 304 клеток / мм 3 ), что аналогично среднему значению. 352 клеток / мм 3 в национальном исследовании.Хотя люди с предшествующими заболеваниями, определяющими СПИД, не соответствовали критериям отбора, примерно у одной пятой испытуемых наблюдались состояния, связанные с ВИЧ, такие как кандидоз кожно-слизистых оболочек, опоясывающий лишай и / или конституциональные симптомы. В нашем исследовании 72% субъектов ранее получали монотерапию зидовудином, тогда как 43% участников исследования ACTG 175 не получали антиретровирусную терапию. За исключением этой разницы, наша подгруппа хорошо представляла базовые демографические данные национального исследования.
В соответствии с результатами, описанными в литературе, 2 полипептидных мотива HSRIG в кодонах 71–75 и 78–82 С-концевой области vpr были строго консервативными.Точно так же цистеин в кодоне 76 и изолейцин в кодоне 83 почти всегда присутствовали. Нуклеотиды в це-спиральном домене выше кодона 70 также хорошо консервативны. Напротив, значительный полиморфизм без доминантного паттерна наблюдался в кодонах 84–86 и кодоне 89. Наиболее важно то, что консенсусный аргинин подтипа B в кодоне 77 был заменен глутамином в 38 (40%) из 96 последовательностей. Эта частота замены R77Q аналогична частоте, обнаруженной в подсчете почти 300 последовательностей подтипа B vpr , зарегистрированных в базе данных Лос-Аламоса до 2002 года.
С-концевые последовательности Vpr не коррелировали с тяжестью предшествующего клинического заболевания. Чтобы определить, может ли замена R77Q или полиморфизм ниже второго мотива HSRIG коррелировать с маркерами ранее существовавшего медленно прогрессирующего заболевания, мы сравнили частоту конкретных изменений С-концевой аминокислоты vpr с исходными абсолютными CD4 + Т-клетками. подсчитывается и с получением либо профилактики пневмонии Pneumocystis carinii, (PCP), либо монотерапии зидовудином до включения в исследование.Никакая замена в кодонах 84–89 не была достаточно распространенной, чтобы оправдать сортировку по клиническому исходу. Абсолютное количество CD4 + Т-клеток при регистрации не коррелировало с наличием мутации глутамина в кодоне 77 и, фактически, противоречило нашей гипотезе. Например, 46% последовательностей от субъектов с исходным абсолютным количеством CD4 + Т-лимфоцитов ниже медианы, 304 клеток / мм 3 , содержали замену R77Q, тогда как это произошло только в 33% последовательностей от субъектов с CD4 * Т-лимфоцитов выше медианы (таблица 1).Аналогичным образом, когда предыдущее лечение рассматривалось как маркер более запущенного и, следовательно, потенциально более интенсивного ранее существовавшего заболевания, 60% субъектов, получавших профилактику PCP, и 41% тех, кто получал монотерапию зидовудином, имели замену R77Q по сравнению с 33% и 37%, соответственно, из тех, кто ранее не лечился (P> 0,25) (таблица 1).
Таблица 1.
Наличие vpr С-концевой замены R77Q в подтипе B, исходные изоляты ВИЧ-1, полученные от исследуемых субъектов, по маркерам клинического статуса и прогрессирования.
Таблица 1.
Наличие vpr С-концевой замены R77Q в подтипе B, исходные изоляты ВИЧ-1, полученные от исследуемых субъектов, по маркерам клинического статуса и прогрессирования.
Мутации на С-конце vpr при включении в исследование также заметно не ослабляли прогрессирование заболевания в ходе исследования. Ни изменение абсолютного количества Т-лимфоцитов CD4 + , ни наклон абсолютного количества Т-лимфоцитов CD4 + за все временные точки исследования не повлияли на изменения в последовательности С-концевой области vpr .В частности, 14 (41%) из 34 субъектов с положительным наклоном абсолютного количества CD4 + Т-клеток и 23 (37%) из 62 человек с отрицательным наклоном имели изменение последовательности в кодоне 77 (таблица 1). Кроме того, с 0 по 128 неделю исходная мутация присутствовала у 44% тех, у кого впоследствии было положительное изменение абсолютного количества CD4 + Т-лимфоцитов, и у 36% тех, у кого впоследствии было отрицательное изменение абсолютного количества CD4 + Т-лимфоцитов, статистически незначимая разница (таблица 1).К сожалению, исследование ACTG 175 было начато до того, как тестирование на вирусную нагрузку плазмы стало обычным явлением, поэтому количественный вирусологический результат не мог быть оценен. В нашем исследовании 28 из 96 субъектов имели клиническую конечную точку, состоящую из СПИД-определяющего или связанного с ВИЧ заболевания или истощения. Из этих 28 человек изоляты из 13 содержали замену R77Q в последовательности vpr , тогда как 25 (37%) из 68 субъектов, не имевших таких конечных точек, содержали ее (таблица 1). Опять же, это статически незначимое (P> .25) тенденция противоречила тому, что можно было бы ожидать, если бы изменение кодона 77 было защитным. Эти протокольные результаты не изменились, когда анализ был стратифицирован по назначению исследуемого препарата.
Обсуждение
Обсуждение . Вариация последовательности участков генома ВИЧ pol, env, gag и nef влияет на клиническое течение заболевания; оценки LTNPs показали C-концевые мутации в дополнительном дополнительном белке, vpr [8, 9, 12–14].В частности, обзор Lum et al. Последовательностей vpr из LTNP, содержащихся в базе данных Лос-Аламоса, показал, что 7% из них демонстрируют полиморфизм в кодоне 77 из vpr по сравнению с 36% таких последовательностей из не-LTNP [9 ]. Такие наблюдения, наряду с предполагаемыми исследованиями in vitro, породили нашу гипотезу о том, что изменения в этой обычно высококонсервативной части генома ВИЧ-1 могут быть связаны с более медленным прогрессированием ВИЧ. В нашей минимально предварительно обработанной группе прогрессоров заболевания ВИЧ-1 объемное секвенирование из архивированных PBMCs показало в 40% образцов изменение согласованной последовательности в кодоне 77 в пределах C-конца vpr. Этот процент намного ниже, чем тот, который ранее был обнаружен в LTNP, но отражает этот процент в последовательностях подтипа B vpr в базе данных Лос-Аламоса.
У наших хорошо охарактеризованных субъектов при регистрации было доступно несколько маркеров, позволяющих различать более продвинутые и менее продвинутые стадии клинического заболевания ВИЧ-1. Однако не было корреляции между интенсивностью предшествующего заболевания, выявленной этими маркерами, и С-концевыми последовательностями vpr субъектов.К сожалению, у нас нет данных о дате заражения ВИЧ-1 у субъектов и, следовательно, о продолжительности их инфицирования до включения в исследование. Конечно, предшествующая профилактика PCP или монотерапия зидовудином или абсолютное количество Т-лимфоцитов CD4 + ниже медианы в исследовании могут частично отражать более длительную продолжительность инфекции; следовательно, наше предположение о том, что эти маркеры исходного статуса отражают либо более вирулентную инфекцию, либо более быстрое прогрессирование заболевания до включения в исследование, необходимо смягчить.Тем не менее, наше предположение может быть в значительной степени справедливым, потому что большинство людей с ВИЧ-1, которые жили в нашем штате на момент проведения нашего исследования, вероятно, были инфицированы в течение ограниченного периода времени в середине 1980-х годов.
Единственное сомнение в том, что наши испытуемые не были полностью терапевтически наивными. В группе ACTG 175 их лечили 1 или 2 нуклеозидными ингибиторами обратной транскриптазы — либо зидовудином, либо диданозином, либо зидовудином в сочетании с диданозином или залцитабином [15].Однако, учитывая, что три четверти из них ранее получали монотерапию зидовудином, по сегодняшним стандартам это были явно минимальные схемы. Таким образом, вызывает разочарование тот факт, что ожидаемая польза от С-концевых мутаций vpr не нашла отражения в маркерах ответа исследования.
Эти результаты предполагают, что ослабляющий болезнь эффект мутации на С-конце vpr , в частности обогащенная замена R77Q, наблюдаемая в большинстве случаев LTNP подтипа B ВИЧ, опубликованных на сегодняшний день, не влияет на течение ВИЧ-инфекции. .Для объяснения этого очевидного противоречия можно привести несколько соображений. Во-первых, возможно, что механизмы R77Q, управляющие ослаблением болезни, также зависят от хозяина, и что мишени-хозяева, с которыми взаимодействует мутант vpr , каким-то образом уникальны для LTNP. Кроме того, наши субъекты участвовали в одном из первых североамериканских испытаний комбинированной антиретровирусной терапии [15], и поэтому наше исследование могло быть смещено в сторону включения субъектов с более агрессивными траекториями болезни.Это возможное «смещение когортной прогрессии» могло затмить относительно слабый эффект против . Наконец, мы выполнили определение последовательности на ампликонах из цельной вирусной ДНК
PBMC. Ограниченные данные предполагают, что квазивиды ВИЧ-1 в С-концевой области vpr могут существовать у одного человека [12]. Если это возможно, то можно ожидать, что наш метод будет недооценивать квазивиды с вариантами меньшинства и склонит наш анализ к обнаружению большего количества вирусов дикого типа.В дополнение к этим ограничениям уместно, что самое крупное опубликованное сравнение последовательностей LTNP и прогрессирующего ВИЧ-1 было получено из базы данных Лос-Аламоса [9], подход, вероятно, искаженный потенциальной систематической ошибкой в собранных последовательностях. Интересно, что консенсусный кодон 77 дикого типа для ВИЧ-1 подтипа C vpr в настоящее время указан как R77Q. По этим причинам мы полагаем, что окончательные ответы относительно клинического значения или отсутствия замены R77Q на С-конце vpr как у LTNP, так и у других инфицированных лиц, требуют дополнительных исследований.
В заключение, хотя данные in vitro свидетельствуют о том, что критические детерминанты апоптоза и остановки клеточного цикла лежат в пределах С-конца vpr, изменения в этой области, очевидно, не повлияли на клинические исходы у пациентов, получавших нуклеозиды в нашем исследовании. По-прежнему может быть, что полиморфизмы vpr, возможно, в сочетании с другими, еще не изученными геномными вариациями ВИЧ-1 или хозяина, действительно играют роль в сохранении статуса LTNP. Тем не менее, дополнительный анализ когорт LTNP и механизмов in vitro должен предшествовать как дальнейшему исследованию образцов из репозиториев естественного происхождения ВИЧ-1, так и началу проспективного испытания на прогрессирующих пациентах, не получавших терапию.
Благодарности
Мы благодарим Джилл Гисбрехт и Мэг Лелонек за помощь в лаборатории и Лилиан Саркинен за подготовку рукописи.
Список литературы
1.,.Сравнение провирусных дополнительных генов у длительно непрогрессоров и прогрессоров инфекции вируса иммунодефицита человека типа 1
,Arch Virol
,2000
, vol.145
(стр.1021
—1027
) 2.,,.Характеристика последовательностей gag и pol от длительно переживших инфекцию вируса иммунодефицита человека типа 1
,Virology
,1998
, vol.240
(стр.36
—49
) 3.,,, Et al.Необычный полиморфизм вируса иммунодефицита человека типа 1, ассоциированный с непрогрессирующей инфекцией
,J Virol
,2000
, vol.74
(стр.4361
—4376
) 4., , , и другие.Анализ вирусных аллелей nef у женщин с непрогрессирующей или медленно прогрессирующей инфекцией ВИЧ-1 [аннотация 88]. Восьмая конференция по ретровирусам и оппортунистическим инфекциям, Чикаго
февраль 2001 г.
5.,,, et al.Иммунологический и вирусологический статус после 14–18 лет инфицирования аттенуированным штаммом ВИЧ-1: отчет группы Sidney Blood Bank Cohort
,N Engl J Med
,1999
, vol.340
(стр.1715
—1722
) 6.,,,.Выделение гена человека, который ингибирует инфекцию ВИЧ-1 и подавляется вирусным белком Vif
,Nature
,2002
, vol.418
(стр.646
—650
) 7.,,,.Изучение биологии вирусного белка ВИЧ-1 R
, ДНКи клеточной биологии
,2002
, vol.21
(стр.679
—688
) 8.,.Митохондриотоксический домен vpr определяет вирулентность ВИЧ-1
,J Clin Invest
,2003
, vol.111
(стр.1455
—1457
) 9.,,, Et al.Vpr R77Q связан с длительной непрогрессирующей ВИЧ-инфекцией и нарушением индукции апоптоза
,J Clin Invest
,2003
, vol.111
(стр.1547
—1554
) 10.,,,,,.Остатки в последовательности HFRIGC ВИЧ-1 vpr , участвующие в активности остановки роста
,Biochem Biophys Res Commun
,1999
, vol.264
(стр.287
—290
) 11.,,.Остатки аргинина на С-конце ВИЧ-1 vpr важны для ядерной локализации и остановки клеточного цикла
,Virology
,1998
, vol.242
(стр.414
—424
) 12.,,, Et al.Генные дефекты, сгруппированные на C-конце гена vpr ВИЧ-1 в длительно не прогрессирующей паре мать и ребенок: эволюция квазивидов vpr in vivo в крови и плазме крови и плазмы
,Virology
,1996
, т.223
(стр.224
—232
) 13.,,, Et al.Дефектные вспомогательные гены у долгосрочного выжившего, инфицированного вирусом иммунодефицита человека типа 1, без излечимого вируса
,J Virol
,1995
, vol.69
(стр.4228
—36
) 14.,,,,.Грубые дефекты генов vpr и vpu ВИЧ типа 1 не могут объяснить различия в количестве копий РНК между длительно бессимптомными и прогрессирующими пациентами
,AIDS Res Hum Retroviruses
,1997
, vol.13
(стр.247
—52
) 15.,,, Et al.Испытание по сравнению монотерапии нуклеозидами с комбинированной терапией у ВИЧ-инфицированных взрослых с числом клеток CD4 от 200 до 500 на кубический миллиметр
,N Engl J Med
,1996
, vol.335
(стр.1081
—90
)© 2004 Американское общество инфекционных болезней
границ | Освещение роли Vpr в ВИЧ-инфекции миелоидных клеток
Введение
Передача половым путем — это преобладающий способ заражения ВИЧ («Global AIDS Update», 2016 г.).В то время как точный тип клеток, нацеленных на ВИЧ в слизистой оболочке половых органов, остается предметом споров (1–3), различные подмножества дендритных клеток (ДК) и макрофагов присутствуют в высоких концентрациях в слизистой оболочке половых органов и, следовательно, могут быть ранними мишенями для заражения. ВИЧ (4–6). Заражение ДК и макрофагами особенно важно, поскольку они обладают уникальной способностью передавать ВИЧ с высокой эффективностью к CD4 + Т-клеткам во время презентации антигена во вторичных лимфоидных органах (5, 7, 8). Как профессиональные антигенпрезентирующие клетки, ДК и макрофаги имеют уникальную клеточную архитектуру, позволяющую инициировать и поддерживать устойчивые взаимодействия с CD4 + Т-клетками.Формирование вирусологического синапса между DC / макрофагами и CD4 + T-клетками обеспечивает направленную доставку ВИЧ к его наиболее благоприятному хозяину: активированным CD4 + T-клеткам (9). Для передачи через слизистые оболочки и создания продуктивной инфекции ВИЧ должен преодолевать не только тканевые барьеры (3), но и ряд присущих клеткам иммунных защитных механизмов или факторов ограничения, таких как APOBEC3G (цитидиндезаминаза, которая резко увеличивает количество мутаций генома. ), тетерин (который предотвращает образование почки вируса ВИЧ и усиливает положительную петлю IFN типа I при подавлении образования вируса) и SAMHD1 (dNTPase, которая ограничивает уровни dNTP в цитоплазме, чтобы препятствовать обратной транскрипции), которые предотвращают лентивирусную инфекцию DC приматов, макрофаги и покоящиеся Т-клетки (10–13).Однако лентивирусы приматов эволюционировали, чтобы противодействовать этим факторам рестрикции, кодируя вспомогательные белки, которые избирательно ингибируют их противовирусную функцию. Хотя предполагается, что он действует в этом качестве, функция Vpr еще предстоит полностью понять.
Vpr, или вирусный белок R, представляет собой белок 14 кДа, кодируемый всеми существующими лентивирусами приматов. Vpr активно упаковывается в вирионы благодаря взаимодействию с областью p 6 Gag (14, 15) и, как таковой, играет роль как на пре-, так и на постинтеграционных стадиях жизненного цикла вируса.Первоначально описанный как вспомогательный белок и, следовательно, незаменимый для репликации вируса in vitro , с тех пор было показано, что он играет важную роль в инфицировании макрофагов и дендритных клеток (16-19). Важно отметить, что функция Vpr необходима для вирусного патогенеза in vivo . В 1993 г. шесть макак-резус были инфицированы патогенным изолятом SIV mac 239, содержащим или не содержащим Vpr (20). В течение 16 недель последовательности Vpr дикого типа выделяли от трех из пяти животных, инфицированных нулевым вирусом Vpr.Более того, у оставшихся двух животных, инфицированных нулевым вирусом Vpr, наблюдался отсроченный патогенез и реверсия к вирусу, экспрессирующему Vpr, к 66 неделям (20, 21). Эти исследования подтвердили ретроспективную работу, которая обнаружила реверсию внутреннего стоп-кодона Vpr в открытую рамку считывания у случайно инфицированного лабораторного работника и экспериментально инфицированных шимпанзе (22, 23). Вместе эти исследования сыграли важную роль в запуске исследований роли Vpr в патогенезе ВИЧ. Наиболее известно, что Vpr вызывает атаксию-телеангиэктазию и Rad3-зависимый (ATR) ответ на повреждение ДНК, или DDR (24).Vpr-опосредованная активация DDR приводит к остановке клеточного цикла от G 2 к M в циклических клетках, что вызывает недоумение, поскольку популяции клеток, инфекции которых, по-видимому, больше всего зависят от присутствия Vpr, окончательно дифференцированы и, следовательно, не подвержены остановке клеточного цикла. . Считается, что способность индуцировать остановку клеточного цикла G 2 в М является преимуществом для вирусной транскрипции, поскольку LTR ВИЧ, как было показано, наиболее активен во время этой фазы клеточного цикла (23, 25). Более того, остатки Vpr, которые придают G 2 способности к остановке клеточного цикла М, находятся под положительным отбором in vivo и, таким образом, являются наиболее хорошо изученными в данной области (26).Остается определить, необходима ли индукция DDR с помощью Vpr, эволюционно законсервированной функции среди всех лентивирусных белков Vpr приматов (27-29), для установления репликации вируса в метаболически покоящихся иммунных клетках-мишенях, таких как моноциты, макрофаги и DCs.
Vpr / Vpx и важность кооптации убиквитин-лигазы DCAF
CRL4 в миелоидных клеткахVpx, или вирусный белок x, возник в результате дупликации Vpr после диверсии лентивирусных клонов приматов, которые дали начало ВИЧ-1 и ВИЧ-2 (30, 31).Vpx играет хорошо изученную роль в деградации ретровирусного рестрикционного фактора SAMHD1 (10, 11). В терминально дифференцированных или нециклирующих клетках SAMHD1 снижает концентрацию dNTPs в цитоплазме, тем самым резко ограничивая обратную транскрипцию (32–35). Vpx связывает SAMHD1 с E3-убиквитинлигазой CUL4A-DDB1 DCAF (DCAF CRL4 ), что приводит к его полиубиквитинированию и протеасомной деградации (10, 11, 36). Большой интерес вызывает идентификация фактора (ов) рестрикции, на который нацелен Vpr, поскольку он аналогичным образом кооптирует комплекс DCAF CRL4 для убиквитинирования белков-мишеней-хозяев.Однако, в отличие от Vpx, инфицирование миелоидных клеток либо ВИЧ-1, либо ВИЧ-2 все еще происходит в отсутствие Vpr, хотя и со значительно разными исходами (19, 24, 37–41). Вероятно, что преимущество репликации, предоставляемое Vpr, заключается в его способности индуцировать DDR, хотя механизмы, с помощью которых Vpr-индуцированная DDR способствует усиленной репликации и распространению вируса in vivo , еще предстоит определить. Множество белков DDR, связанных с комплексом Vpr-DCAF CRL4 (24, 37–39, 41), предполагает, что Vpr, кооптируя комплекс белков-хозяев, участвующих во множестве клеточных путей, смог максимизировать свое влияние на интерфейс вируса и хозяина для содействия распространению ВИЧ.
Остатки Vpr, задействованные в DCAF
CRL4 ЗадержкаЯМР-структура ВИЧ-1 Vpr дает представление о том, как он взаимодействует с множеством белков. Оба N- и C-конца неструктурированы (нуклеотиды 1–16 и 77–96 соответственно) и фланкируют три α-спирали от нуклеотидов 17–33, 38–50 и 56–77 (42). Vpr ВИЧ-2, а также близкородственные аллели Vpr из SIV smm и SIV mac , как предполагается, структурно гомологичны таковому из ВИЧ-1.В то время как неструктурированные C- и N-концевые домены способствуют взаимодействию с мишенями-хозяевами, DCAF CRL4 — связывающий домен изолирован от третьей α-спирали (42–45). Например, мутанты Vpr ВИЧ-1 Q65R и H71R и соответствующие остатки в аллелях Vpr ВИЧ-2 / SIV mac попадают в эту область и полностью исключают взаимодействия Vpr-DCAF CRL4 . Эти мутации предотвращают опосредованное Vpr усиление транскрипции в MDDC (19), уменьшают деградацию белков ответа на множественные повреждения ДНК (46-50) и предотвращают остановку клеточного цикла G 2 / M в CD4 + Т-клетках (51).Кроме того, устранение взаимодействия Vpr-DCAF CRL4 , как это происходит с мутацией VprQ65R (хотя и не с мутацией VprQ77R), было связано с длительным непрогрессированием in vivo (52). Исследования опосредованного DCAF CRL4 усиления инфекции в миелоидных клетках используют эти избранные мутации для определения механизмов действия и, таким образом, заслуживают упоминания.
DCAF
CRL4 -Зависимые роли Vpr в миелоидных клеткахБелки ответа на повреждение ДНК
Урацил ДНК-гликозилаза человека
Макрофаги, происходящие из моноцитов (MDM), имеют высокое соотношение dUTP / TTP в своей цитоплазме, что может привести к неправильному включению урацила в геном с обратной транскрипцией.Было обнаружено, что соотношение dUTP / TTP в макрофагах достигает 60 (53, 54). Урацил-ДНК-гликозилаза человека (hUNG) удаляет неправильно включенный UTP и задействует дополнительные ферменты репарации в месте мутации генома. Таким образом, было выдвинуто предположение, что опосредованное Vpr ВИЧ-1 DCAF CRL4 убиквитинирование и протеасомная деградация hUNG ограничивают репликацию вируса либо за счет деградации урацилированной вирусной ДНК до интеграции, либо посредством транскрипционного вмешательства урацилированного провируса (53, 55, 56). ).Однако из-за изначально низких уровней hUNG в MDM (56) полезность урацил-зависимого ограничения ВИЧ-1 в MDM ограничена. Более того, инфицирование MDDCs ВИЧ-1, экспрессирующим hUNG-связывающий дефицитный мутант VprW54R, не приводит ни к ослаблению транскрипции, ни к дефициту вирусного распространения (19). Таким образом, значение деградации hUNG комплексом Vpr-DCAF CRL4 (39, 57) остается неясным.
SLX4-SLX / MUS81-EME1
Важность Vpr в пути репарации соединений Холлидея представляет большой интерес, поскольку обещает обеспечить понимание роли Vpr на стадии вирусной интеграции.Исходные сообщения предполагают, что ВИЧ-1 Vpr-DCAF CRL4 -опосредованное убиквитинирование MUS81, которое в присутствии фосфорилированного EME1 и киназно-активного PLK1 преждевременно активирует четвертичный эндонуклеазный комплекс SLX4com (47). Было показано, что эта активация предшествует остановке клеточного цикла G 2 / M и приводит к образованию фокусов FANCD2 в результате активации пути анемии Фанкони (47). В частности, было показано, что связанная с вирионом Vpr-опосредованная активация SLX4com предотвращает продукцию IFN типа I, что имеет большое значение из-за множества генов, стимулированных интерфероном (ISG), которые модулируют функцию миелоидных клеток и определяют эффективность распространения вируса через хозяина. (47).Однако последующие исследования показали только Vpr-DCAF CRL4 зависимую деградацию SLX4com субъединиц MUS81-EME1 (50, 58-60) и не выяснили, подавляет ли активный SLX4com детектирование ВИЧ-1 в миелоидных клетках врожденным иммунитетом. Кроме того, взаимодействие SLX4com с Vpr не консервативно среди всех лентивирусных аллелей Vpr приматов (59). В совокупности опубликованные на данный момент результаты предполагают, что Vpr-опосредованная активация SLX4com не играет консервативной роли в подавлении врожденного иммунного обнаружения лентивирусов приматов в миелоидных клетках.
Геликазоподобный фактор транскрипции (HLTF)
Геликазоподобный фактор транскрипции, или HLTF, является мишенью Vpr-опосредованной деградации DCAF CRL4 (46, 48). Подобно UNG2 и SLX4-SLX1 / MUS81-EME1, HLTF участвует в репарации повреждений ДНК. В частности, HLTF имеет решающее значение для ремоделирования и восстановления застопорившихся репликационных вилок (61). Хотя HLTF расщепляется в макрофагах зависимым от Vpr-DCAF CRL4 образом, неясно, противодействует ли HLTF репликации вируса в миелоидных клетках.
Экзонуклеаза 1
Экзонуклеаза 1, или Exo1, представляет собой 5′-3′-экзонуклеазу Rad2 / XPG, участвующую во многих процессах репарации ДНК, что обеспечивает стабильность генома на протяжении всего клеточного цикла (62). Exo1 недавно был идентифицирован как субстрат для полиубиквитинирования и протеасомной деградации Vpr-DCAF CRL4 в CD4 + Т-клетках (49). Авт. Предполагают, что антагонизм Exo1 предотвращает вредный процессинг обратной транскрипции и промежуточных продуктов вирусной интеграции и тем самым приписывает ограничение Exo1 ассоциированному с вирионом Vpr скорее, чем его de novo синтезируемому партнеру (49).На данный момент не показано, что Exo1 играет роль в развитии ВИЧ-1 инфекции макрофагов или дендритных клеток.
Хотя эти опубликованные исследования подчеркивают многочисленные взаимодействия Vpr с различными белками DDR, вклад этих взаимодействий в вирусный патогенез остается неясным. Несмотря на то, что в случае ВИЧ-1-инфекции мало изучены, существует надежная литература, связывающая передачу сигналов врожденного иммунитета и DDR (63, 64). Следует отметить, что манипуляции с DDR не уникальны для ВИЧ.Скорее, это общая патогенная стратегия, широко используемая на взаимодействии хозяев с бактериями и вирусами, которые могут способствовать репликации патогенов и патогенезу (65). Вирус герпеса саркомы Капоши, например, кодирует белок (связанный с латентностью ядерный антиген или LANA), который изолирует Rad50, Mre11 и NBS1, все члены комплекса MRN активатора передачи сигналов DDR, чтобы предотвратить цитоплазматическое зондирование вирусной ДНК и активацию врожденного иммунитета (66 ). Другой пример вирусного нарушения пути DDR включает мышиный γ-герпесвирус, который специфически кодирует orf36, роль которого заключается в индукции ATM-зависимого фосфорилирования DDR и h3AX (67).В отсутствие активации orf36 или ATM репликация вируса ослабляется, что указывает на роль DDR в облегчении репликации вируса (67). В целом очевидно, что Vpr использует DCAF CRL4 для индукции DDR с потенциально разными результатами в разных популяциях клеток. Что остается неясным, так это то, как активация DDR и взаимодействие Vpr с белками репарации ДНК позволяет вирусам уклоняться от иммунного обнаружения в миелоидных клетках. Поскольку кинетика обратной транскрипции ВИЧ-1 в миелоидных клетках относительно медленная, есть соблазн предположить, что манипулирование различными путями DDR является консервативной стратегией лентивирусных аллелей Vpr приматов для преодоления преждевременной репарации вирусных промежуточных продуктов обратной транскрипции хозяином (63). , хотя окончательные доказательства этой гипотезы отсутствуют.Скорее всего, активация DDR способствует нескольким дискретным стадиям жизненного цикла вируса. Например, Vpr может индуцировать DDR как через пути ATM, так и через ATR (24, 68). Неразрешенная активность ATM может привести к активации NF-κB (69) и увеличению выработки воспалительных цитокинов, таких как IL-6, оба из которых могут привести к усиленной экспрессии вирусных генов и макрофагозависимой передаче ВИЧ-1 к CD4 + T-клеткам ( 70).
Функции Vpr в дерепрессии транскрипции
Улучшение транскрипции
Предыдущая работа нашей группы показала постинтеграционный дефект в дендритных клетках, происходящих из моноцитов (MDDC), инфицированных Vpr-дефицитным ВИЧ-1 (19).Инфекции в отсутствие Vpr, ассоциированного с вирионом, характеризовались низкой провирусной транскрипцией, несмотря на аналогичные уровни интеграции, и снижением инфицирования CD4 + Т-клеток в совместных культурах (19). Этот дефект зависит от связывания Vpr с DCAF CRL4 , поскольку он полностью устраняется при инфицировании мутантами Vpr (Q65R или H71R), лишенными взаимодействий DCAF CRL4 . Следует отметить, что многие вирусы, помимо ВИЧ-1, в первую очередь вирус гепатита B, также могут манипулировать убиквитинлигазой E3 DCAF CRL4 для усиления транскрипции (71).Хотя механизм Vpr-опосредованного усиления транскрипции ВИЧ-1 остается неясным, предыдущие исследования показали, что Vpr-опосредованная деградация HDAC (38) и членов комплекса ремоделирования хроматина NuRD (72) может усиливать транскрипцию в глобальном масштабе. Кроме того, DCAF CRL4 также играет хорошо известную роль в деградации репрессора транскрипции, ATF3, который необходим для коррекции УФ-повреждений (73). Это объяснение не является удовлетворительным, учитывая зависимость усиления транскрипции от типа клеток.Разложен или изолирован специфичный для MDDC репрессор / активатор, остается неизвестным и требует дальнейшего исследования.
ТЕТ2
Недавно было показано, что члены семейства TET ДНК-диоксигеназ разлагаются зависимым образом от Vpr-DCAF CRL4 (70). В миелоидных клетках TET2 естественно моноубиквитинируется. N-терминальное моноубиквитилирование TET2 делает возможным эффективное связывание с хроматином и последующее задействование аппарата ремоделирования хроматина и факторов транскрипции.Однако в присутствии Vpr TET2 быстро полиубиквитинируется в сайте, независимом от его природного сайта моноубиквитилирования, и подвергается DCAF CRL4 -зависимой протеасомной деградации (70). Это актуально для миелоидных клеток, поскольку TET2 является вышестоящим супрессором экспрессии IL-6. TET2 рекрутирует HDAC1 и HDAC2 на промотор IL-6, тем самым подавляя транскрипцию IL-6. Важно отметить, что в макрофагах, происходящих из моноцитов, и в линии моноцитарных клеток THP-1 отсутствие Vpr-опосредованной деградации TET2 было связано со сниженным высвобождением вирусных частиц и более медленным распространением инфекции ВИЧ-1.После нокаута TET2 различия в инфекции между Vpr-компетентными и Vpr-дефицитными вирусами были потеряны. Поскольку IL-6 долгое время считался усилителем транскрипции ВИЧ в моноцитах (70, 74–76), эти данные дополнительно наводят на мысль о прямой связи между Vpr, деградацией TET2 и устойчивым продуцированием IL-6, что может привести к повышенная эффективность распространения вируса от миелоидных клеток к CD4 + Т-клеткам.
Эпигенетическая регуляция провируса
До недавнего времени не было известно, существуют ли внутренние механизмы хозяина для ограничения ретровирусной репликации после интеграции.Однако недавние исследования идентифицировали новый метод внутриклеточного ограничения: метод отложения меток метилирования, подавляющего транскрипцию, на провирусных LTR. После обратной транскрипции и интеграции LTR провирусов в гетерохроматине метилируются посредством последовательного рекрутирования HP1, а метилтрансфераза Suv39h2 (77). Триметилированный h4K9 рекрутирует комплекс HUSH (HUman Silencing Hub), состоящий из трех компонентов; ТАСОР, МПП8 и перифилин (78).Хотя комплекс HUSH сам по себе не обладает активностью метилтрансферазы, HUSH рекрутирует метилазу SETDB1, которая индуцирует дальнейшее метилирование h4K9me3 провируса. Примечательно, что shRNA-опосредованный нокдаун каждого сложного белка HUSH спасает экспрессию эндогенного и экзогенного ретровирусного гена, тем самым показывая важность его четвертичной сборки для репрессии транскрипции (78). Интересно, что TASOR нацелен клонов Vpx SIVmac / HIV-2 для DCAF CRL4 -опосредованного полиубиквитинирования и протеасомной деградации, тем самым увеличивая транскрипционную активность провирусов, которая в противном случае была бы подавлена (78-80).В то время как комплекс HUSH может репрессировать транскрипцию из интегрированного LTR ВИЧ-1 (78, 80, 81), удивительно, что Vpr ВИЧ-1 не нацелен на белки комплекса HUSH для деградации (79). Однако было показано, что множественные аллели Vpr от лентивирусов предков приматов к ВИЧ-1, включая аллели, происходящие от SIV AGM , SIV MUS2 и SIV SAB , предотвращают опосредованное HUSH молчание (79, 80). Эти исследования знаменуют начало исследований антагонизма Vpx / Vpr противовирусных белков хозяина на уровне провирусной ДНК.Хотя HUSH-опосредованное подавление транскрипции не является специфическим для миелоида противовирусным механизмом, комплекс HUSH и связанные с ним фасилитаторы активны в миелоидных клонах. Недавние исследования в литературе предоставляют доказательства эпигенетического контроля провоспалительных цитокиновых ответов в макрофагах (82, 83). Например, гистоновые метилтрансферазы, SETDB1 и Smyd2, сильно подавляют TLR4-опосредованную индукцию продукции IL-6 и TNFα, а мыши с макрофаг-специфическим дефицитом SetDB1 гиперчувствительны к провокации эндотоксином (82).Хотя антагонизм комплекса HUSH не был приписан Vpr ВИЧ-1, подавление транскрипции LTR ВИЧ-1 в MDDC в отсутствие Vpr (19) предполагает существование дополнительных механизмов внутренней репрессии транскрипции миелоидными клетками, на которые нацелены ВИЧ-1 Впр.
Дайсер и процессинг miRNA
Модуляция путей РНК-интерференции (RNAi) и микроРНК (miRNA) является неотъемлемым средством, с помощью которого патогены узурпируют функции хозяина в свою пользу (84, 85).В частности, давно известно, что микроРНК играют роль в репликации ВИЧ во многих популяциях клеток. Например, miR-29a участвует в подавлении уровней мРНК ВИЧ посредством связывания с 3′-UTR РНК ВИЧ и последующего прикрепления к белкам P-тельца и комплексам RISC (86). Дайсер необходим для обработки субстратов пре-miRNA, чтобы выявить двухцепочечный комплекс miRNA, который затем связывается с комплексом RISC и подавляет экспрессию целевой мРНК либо посредством ингибирования трансляции, либо посредством деградации мРНК (87).Недавние исследования также продемонстрировали способность miRNA отрицательно регулировать провоспалительные реакции в макрофагах путем ограничения ремоделирования хроматина и усиления транскрипционного молчания промоторов выбранных воспалительных генов (88). Интересно, что Dicer был идентифицирован в комплексе с Vpr-DCAF CRL4 до его деградации и истощения Dicer в инфицированных MDM, как было показано, увеличивает репликацию вируса с помощью неизвестных механизмов (89). Мы утверждаем, что Vpr-Dicer-зависимая модуляция экспрессии выбранной miRNA может вносить вклад в дерепрессию воспалительных реакций.Следует отметить, что Vpr-опосредованное истощение Dicer также было показано в CD4 + Т-клетках и, как таковое, не является миелоид-специфическим антагонистом врожденной рестрикции (89). Роль деградации Dicer еще предстоит полностью понять, особенно в связи с расширением исследований функции miRNA и RNAi в патогенезе ВИЧ (90).
DCAF
CRL4 -Независимые роли Vpr в миелоидных клеткахОбращение с конвертами
Миелоидные клетки часто населяют ткани слизистых оболочек и, как таковые, способны распространять ВИЧ с периферии к участкам, где в большом количестве находятся активированные CD4 + Т-клетки.Макрофаги и дендритные клетки способны переносить вирус через цис- или транс-инфекцию. Оба метода способствуют концентрации вирионов в инфекционном синапсе и тем самым значительно увеличивают вероятность заражения CD4 + Т-клетками (9, 91, 92). Два исследования изучали роль Vpr в концентрации и доставке вируса в вирусологический синапс. Оба этих исследования показывают, что в отсутствие Vpr, Env-положительные вирионы доставляются в лизосомы для деградации, тем самым снижая эффективность распространения макрофагов для CD4 + T-клеточного вируса при низкой множественности инфекции (93, 94).Напротив, наши исследования того, способствует ли Vpr уклонению от этого Env-зависимого снижения высвобождения вируса в других типах клеток, таких как MDDC, не дали аналогичных результатов (19), предполагая, что влияние Vpr на экспрессию Env может быть ограничено до специфические типы клеток и не всегда наблюдаются.
IFN типа I и провоспалительные реакции
Появляется все больше свидетельств того, что Vpr модулирует иммунный ответ миелоидных клеток, способствуя репликации и распространению вируса по всему хозяину.Ранние исследования предполагали возможный дефект активации MDM и MDDC при лечении рекомбинантным Vpr (95). Этот дефект характеризовался низкой экспрессией CD33 на поверхности, плохой активацией CD80 / 86 и нарушенной презентацией антигена активированным CD4 + Т-клеткам (95). В отличие от исследований, в которых использовалось экзогенное добавление рекомбинантного Vpr, преобладали исследования, изучающие роль Vpr в контексте вирусной инфекции. Например, de novo экспрессия Vpr в продуктивно инфицированных MDDC индуцировала продукцию провоспалительных цитокинов (TNF-α, IL-6 и IL-8) (70, 96, 97).Предыдущая работа нашей группы показала усиление провирусной транскрипции в MDDC в присутствии Vpr (19). Другие исследования аналогичным образом показали роль Vpr в провирусной транскрипции. Лю и др. показали, что Vpr изменяет доступность гетеродимера NF-κB p50-p65 и AP1 (98), оба из которых необходимы для инициации транскрипции ВИЧ с 5′-LTR (99, 100) и экспрессии провоспалительных цитокинов. В этом исследовании было показано, что Vpr облегчает полиубиквитинирование и последующее фосфорилирование (активацию) TAK1, вышестоящего регулятора NF-κB и AP1 (98).Прерывание фосфорилирования TAK1 и, таким образом, ингибирование его активации, значительно снижает провирусную транскрипцию (98).
Как наше исследование (19), так и работа, показывающая Vpr-опосредованную модификацию TAK1 (98), важны в свете недавней идентификации нового пути обнаружения вируса: такого, в котором хозяин зондирования de novo экспрессирует интрон-содержащий ВИЧ. -1 РНК (icRNA ВИЧ) в MDM и MDDC приводит к экспрессии ISG и продукции провоспалительных хемокинов и цитокинов, включая IP-10 и IL-15 (101).IP-10 представляет собой воспалительный хемокин и лиганд рецептора CXCR3 (102), в то время как IL-15 является цитокином γ-цепи, критическим для пролиферации и гомеостаза Т-клеток (103). CXCR3 экспрессируется на активированных CD4 + Т-клетках, и, таким образом, секреция IP-10 продуктивно инфицированными миелоидными клетками может привести к дополнительному привлечению чувствительных к вирусу клеток в сайты репликации вируса в периферических тканях. Кроме того, воздействие IL-15 может приводить к фосфорилированию SAMHD1 и инактивации его активности dNTPase, тем самым снимая ограничения на репликацию вируса в покоящихся CD4 + Т-клетках (104).Интересно, что ингибитор TAK1 снижает продукцию IP-10 из макрофагов, инфицированных ВИЧ-1 (101). Таким образом, в этой модели Vpr-зависимая усиленная провирусная транскрипция потенциально увеличивает пул вирусных icRNA, подверженных внутреннему иммунному восприятию клетки, что приводит к установлению провоспалительного состояния и усиленному распространению вируса.
Следует отметить, что увеличение вирусной транскрипции в миелоидных клетках является палкой о двух концах для ВИЧ, поскольку также происходит индукция экспрессии ISG и установление предполагаемого антивирусного состояния.Хотя механизм восприятия нуклеиновой кислоты, ответственный за обнаружение icRNA ВИЧ, еще не определен, Vpr может блокировать фосфорилирование и ядерную транслокацию фактора регуляции интерферона 3 (IRF3), индуцируя убиквитинирование и протеосомную деградацию IRF3, хотя это было показано только для CD4 +. Линии Т-клеток (105). Кроме того, было показано, что Vpr связывается с TBK1 в миелоидных клетках и предотвращает его фосфорилирование, тем самым предотвращая индукцию продукции IFN типа I (106). Таким образом, мы утверждаем, что Vpr может действовать, способствуя NF-κB-зависимым провоспалительным ответам, одновременно способствуя подавлению индукции противовирусной защиты хозяина.Более того, индукция ISG, таких как CD169, в MDM и MDDC при восприятии хозяином icRNA ВИЧ в миелоидных клетках (101) может еще больше склонить чашу весов к усиленному распространению вируса, а не к ограничению вируса. Например, индуцированная экспрессия CD169 на ВИЧ-инфицированных макрофагах и дендритных клетках может способствовать передаче CD4 + Т-клеток от клетки к клетке через инфекционные синапсы (101, 107, 108). Вместе эти исследования указывают на роль Vpr как белка, который тщательно управляет множеством вирусных сенсорных систем, чтобы вызвать рекрутирование дополнительных клеточных мишеней вируса, избегая при этом противовирусного иммунитета.
Заключение
Очевидно, что Vpr играет важную роль в инфицировании миелоидных клеток (см. Рисунок 1). Ряд тканевых макрофагов, таких как микроглия, клетки Купфера, альвеолярные, кишечные, тестикулярные и вагинальные макрофаги, содержат провирусную ДНК (109–113), а тканевые макрофаги, по оценкам, компрометируют до 4% инфицированных клеток в vivo (114) и, что важно, может оставаться устойчиво инфицированным ВИЧ-1 даже в присутствии АРТ (109–112, 115).Возможно, что Vpr-опосредованная DDR активирует провоспалительное состояние, которое способствует созданию резидентного в тканях резервуара миелоидных клеток, в результате чего вирус эффективно распространяется благодаря устойчивому производству вирионов и усилению межклеточных контактов между ВИЧ-инфицированными миелоидными клетками. клетки и CD4 + Т-клетки. Таким образом, инфекция миелоидных клеток является мостом между относительно враждебными участками заражения вирусом (в первую очередь, периферическими тканями слизистой оболочки) и ключевой мишенью ВИЧ; CD4 + Т-клетки.Кажется вероятным, что истинная ценность Vpr in vivo заключается в его универсальности, позволяющей избежать вирусного ограничения как до, так и после интеграции в миелоидные клетки. В будущих исследованиях необходимо будет рассмотреть относительную важность каждой из известных функций Vpr in vivo .
Рисунок 1 . Роль Vpr в инфицировании миелоидных клеток. Краткое изложение многочисленных функций, которые Vpr играет в миелоидных клетках, от усиления транскрипции и индукции DDR до секреции провоспалительных цитокинов.Красным цветом показаны процессы, в которых Vpr был непосредственно исследован.
Взносы авторов
Все перечисленные авторы внесли существенный, прямой и интеллектуальный вклад в работу и одобрили ее к публикации.
Финансирование
Эта работа была поддержана грантами NIH R01AI064099, P30AI042853 и R01AG060890 (SG).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
2. Баррето-де-Соуза В., Аракелян А., Марголис Л., Ванпуй К. Передача ВИЧ-1 вагинальным путем: внеклеточный или связанный с клетками вирус? Am J Reproduc Immunol. (2014) 71: 589–99. DOI: 10.1111 / aji.12240
CrossRef Полный текст | Google Scholar
3. Бургенер А., Макгоуэн И., Клатт Н.Р. Взаимодействие ВИЧ и слизистого барьера: последствия для передачи и патогенеза. Курр Опин Иммунол . (2015) 36: 22–30. DOI: 10.1016 / j.coi.2015.06.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
4. Пена-Круз В., Агосто Л.М., Акияма Х., Олсон А., Моро И., Ларрие Дж. Р. и др. ВИЧ-1 реплицируется и сохраняется в эпителиальных дендритных клетках влагалища. Дж. Клин Инвест . (2018) 128: 3439–44. DOI: 10.1172 / JCI98943
PubMed Аннотация | CrossRef Полный текст | Google Scholar
5. Родригес-Гарсия М., Шен З., Барр Ф. Д., Бош А. В., Акерман М. Е., Каппес Дж. К. и др. Дендритные клетки женского репродуктивного тракта человека быстро захватывают ВИЧ и реагируют на него. Иммунол слизистой оболочки . (2017) 10: 531–44. DOI: 10,1038 / mi.2016.72
PubMed Аннотация | CrossRef Полный текст | Google Scholar
7. Peressin M, Proust A, Schmidt S, Su B, Lambotin M, Biedma ME, et al. Эффективный перенос ВИЧ-1 в транс- и цис-форме от дендритных клеток Лангерганса и макрофагов к аутологичным Т-лимфоцитам. СПИД . (2014) 28: 667–77. DOI: 10.1097 / QAD.0000000000000193
PubMed Аннотация | CrossRef Полный текст | Google Scholar
8.Турвилл С.Г., Сантос Дж. Дж., Фрэнк И., Кэмерон ПУ, Уилкинсон Дж., Миранда-Саксена М. и др. Поглощение вируса иммунодефицита, оборот и двухфазный перенос в дендритных клетках человека. Кровь . (2004) 103: 2170–9. DOI: 10.1182 / кровь-2003-09-3129
PubMed Аннотация | CrossRef Полный текст | Google Scholar
10. Грецка К., Хао С., Гершевска М., Суонсон С.К., Кесик-Бродацка М., Шривастава С. и др. Vpx снимает подавление ВИЧ-1 инфекции макрофагов, опосредованной белком SAMHD1. Природа . (2011) 474: 658–61. DOI: 10.1038 / nature10195
PubMed Аннотация | CrossRef Полный текст | Google Scholar
11. Laguette N, Sobhian B., Casartelli N, Ringeard M, Chable-Bessia C., Ségéral E, et al. SAMHD1 представляет собой специфический для дендритных и миелоидных клеток фактор рестрикции ВИЧ-1, которому противодействует Vpx. Природа . (2011) 474: 654–7. DOI: 10.1038 / природа10117
PubMed Аннотация | CrossRef Полный текст | Google Scholar
13. Sheehy AM, Gaddis NC, Choi JD, Malim MH.Выделение человеческого гена, который подавляет инфекцию ВИЧ-1 и подавляется вирусным белком Vif. Природа . (2002) 418: 646–50. DOI: 10.1038 / nature00939
PubMed Аннотация | CrossRef Полный текст | Google Scholar
14. Селиг Л., Пейдж Дж. К., Танчу В., Превераль С., Берлиоз-Торрент С., Лю LX и др. Взаимодействие с доменом p6 предшественника gag опосредует включение в вирионы белков Vpr и Vpx из лентивирусов приматов. Дж Вирол . (1999) 73: 592–600.
PubMed Аннотация | Google Scholar
15. Пакстон В., Коннор Р.И., Ландау Н.Р. Включение Vpr в вирионы вируса иммунодефицита человека типа 1: потребность в области p6 gag и мутационного анализа. J Virol. (1993) 67: 7229–37.
PubMed Аннотация | Google Scholar
16. Кэмпбелл Б.Дж., Хирш В.М. Vpr вируса обезьяньего иммунодефицита африканских зеленых мартышек необходим для репликации в макрофагах и лимфоцитах макак. Дж Вирол .(1997) 71: 5593–602.
PubMed Аннотация | Google Scholar
17. Коннор Р.И., Чен Б.К., Чхве С., Ландау Н.Р. Vpr необходим для эффективной репликации вируса иммунодефицита человека типа 1 в мононуклеарных фагоцитах. Вирусология . (1995) 206: 935–44. DOI: 10.1006 / viro.1995.1016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
18. Экштейн Д.А., Шерман М.П., Пенн М.Л., Чин П.С., Грин В.С., Голдсмит М.А. Vpr ВИЧ-1 увеличивает вирусную нагрузку, облегчая инфицирование тканевых макрофагов, но не деля CD4 + Т-клетки. J Exp Med . (2001) 194: 1407–19. DOI: 10.1084 / jem.194.10.1407
CrossRef Полный текст | Google Scholar
19. Миллер С.М., Акияма Х., Агосто Л.М., Эмери А., Эттингер С.Р., Сванстром Р.И. и др. Связанный с вирионом Vpr снижает постинтеграционную блокаду ВИЧ-1 инфекции дендритных клеток. Дж Вирол . (2017) 91: e00051-17. DOI: 10.1128 / JVI.00051-17
PubMed Аннотация | CrossRef Полный текст | Google Scholar
20. Ланг SMM, Вигер С., Шталь-Хенниг С., Кулибали Дж., Хунсманн Дж., Мюллер Х. и др.Важность Vpr для заражения макак-резусов вирусом иммунодефицита обезьян. Дж Вирол . (1993) 67: 902–12.
PubMed Аннотация | Google Scholar
21. Hoch J SM, Lang M, Weeger C, Stahl-Hennig C., Coulibaly U, Dittmer G, et al. Мутант с делецией vpr вируса иммунодефицита обезьян вызывает СПИД у макак-резусов. Дж Вирол . (1995) 69: 4807–13.
PubMed Аннотация | Google Scholar
22. Бомонт Т., ван Нуэнен А., Броерсен С., Блаттнер В.А., Лукашов В.В., Шуйтмейкер Х.Обращение вируса иммунодефицита человека типа 1 IIIB к фенотипу, устойчивому к нейтрализации, у случайно инфицированного лабораторного работника с прогрессирующим клиническим течением. Дж Вирол . (2001) 75: 2246–52. DOI: 10.1128 / JVI.75.5.2246-2252.2001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
23. Goh WC, Rogel ME, Kinsey CM, Michael SF, Fultz PN, Nowak MA, et al. Vpr ВИЧ-1 увеличивает экспрессию вируса путем манипулирования клеточным циклом: механизм отбора Vpr in vivo . Нат Мед . (1998) 4:65. DOI: 10,1038 / нм0198-065
PubMed Аннотация | CrossRef Полный текст | Google Scholar
24. Рошаль М., Ким Б., Чжу И., Нгием П., Планеллес В. Активация ATR-опосредованного ответа на повреждение ДНК вирусным белком ВИЧ-1 R. J Biol Chem . (2003) 278: 25879–86. DOI: 10.1074 / jbc.M303948200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
25. Гуммулуру С., Эмерман М. Клеточный цикл и Vpr-опосредованная регуляция экспрессии вируса иммунодефицита человека типа 1 в первичных и трансформированных Т-клеточных линиях. Дж Вирол . (1999) 73: 5422–30.
PubMed Аннотация | Google Scholar
26. Yedavalli VR, Chappey C, Ahmad N. Сохранение интактного гена vpr вируса иммунодефицита человека типа 1 после передачи от матери ребенку. Дж Вирол . (1998) 72: 6937–43.
PubMed Аннотация | Google Scholar
28. Planelles V, Jowett JB, Li QX, Xie Y, Hahn B., Chen IS. Vpr-индуцированная остановка клеточного цикла консервативна среди лентивирусов приматов. Дж Вирол .(1996) 70: 2516–24.
PubMed Аннотация | Google Scholar
29. Stivahtis GL, Soares MA, Vodicka MA, Hahn BH, Emerman M. Сохранение и специфичность хозяина Vpr-опосредованной остановки клеточного цикла предполагают фундаментальную роль в эволюции и биологии лентивируса приматов. Дж Вирол . (1997) 71: 4331–8.
PubMed Аннотация | Google Scholar
30. Фрегосо О.И., Ан Дж., Ван С., Меренс Дж., Сковронски Дж., Эмерман М. Эволюционное переключение специфичности Vpx / Vpr приводит к дивергентному распознаванию рестрикционного фактора SAMHD1. PLoS Патог . (2013) 9: e1003496. DOI: 10.1371 / journal.ppat.1003496
PubMed Аннотация | CrossRef Полный текст | Google Scholar
31. Лим ES, Fregoso OI, McCoy CO, Matsen FA, Malik HS, Emerman M. Способность лентивирусов приматов разрушать фактор рестрикции моноцитов SAMHD1 предшествовала рождению вирусного вспомогательного белка Vpx. Клеточный микроб-хозяин . (2012) 11: 194–204. DOI: 10.1016 / j.chom.2012.01.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
32.Baldauf HM, Pan X, Erikson E, Schmidt S, Daddacha W., Burggraf M, et al. SAMHD1 ограничивает инфекцию ВИЧ-1 в покоящихся CD4 (+) Т-клетках. Нат Мед . (2012) 18: 1682–7. DOI: 10,1038 / нм.2964
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33. Гужон С., Арфи В., Пертел Т., Любан Дж., Лиенард Дж., Ригал Д. и др. Характеристика функции Vpx вируса иммунодефицита обезьян SIVSM / вируса иммунодефицита человека 2 типа в миелоидных клетках человека. Дж Вирол . (2008) 82: 12335–45.DOI: 10.1128 / JVI.01181-08
PubMed Аннотация | CrossRef Полный текст | Google Scholar
34. Лахуасса Х., Даддача В., Хофманн Х., Айинде Д., Лог Э. К., Драгин Л. и др. SAMHD1 ограничивает репликацию вируса иммунодефицита человека типа 1, истощая внутриклеточный пул дезоксинуклеозидтрифосфатов. Нат Иммунол . (2012) 13: 223–8. DOI: 10.1038 / ni.2236
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35. Descours B, Cribier A, Chable-Bessia C, Ayinde D, Rice G, Crow Y и др.SAMHD1 ограничивает обратную транскрипцию ВИЧ-1 в покоящихся CD4 (+) Т-клетках, Retrovirology . (2012) 9:87. DOI: 10.1186 / 1742-4690-9-87
PubMed Аннотация | CrossRef Полный текст | Google Scholar
36. Hofmann H, Logue EC, Bloch N, Daddacha W., Polsky SB, Schultz ML, et al. Дополнительный лентивирусный белок Vpx нацелен на деградацию SAMHD1 в ядре. Дж Вирол . (2012) 86: 12552–60. DOI: 10.1128 / JVI.01657-12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
37.Белзил Дж. П., Ричард Дж., Ружо Н., Сяо Ю., Коэн Э. А.. Vpr ВИЧ-1 индуцирует K48-связанное полиубиквитинирование и протеасомную деградацию целевых клеточных белков, чтобы активировать ATR и способствовать остановке G2. Дж Вирол . (2010) 84: 3320–30. DOI: 10.1128 / JVI.02590-09
PubMed Аннотация | CrossRef Полный текст | Google Scholar
38. Romani B, Baygloo NS, Hamidi-Fard M, Aghasadeghi MR, Allahbakhshi E. Белок Vpr ВИЧ-1 индуцирует протеасомную деградацию ассоциированных с хроматином HDAC класса I для преодоления латентной инфекции макрофагов. Дж. Биол. Хим. . (2016) 291: 2696–711. DOI: 10.1074 / jbc.M115.689018
PubMed Аннотация | CrossRef Полный текст | Google Scholar
39. Schröfelbauer B, Yu Q, Zeitlin SG, Landau NR. Vpr вируса иммунодефицита человека типа 1 индуцирует деградацию урацил-ДНК гликозилаз UNG и SMUG. Дж Вирол . (2005) 79: 10978–87. DOI: 10.1128 / JVI.79.17.10978-10987.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40. Уэно Ф., Шиота Х., Мияура М., Йошида А., Сакураи А., Тацуки Дж. И др.Белки Vpx и Vpr ВИЧ-2 регулируют вирусную инфекционность посредством особого механизма в лимфоцитарных клетках. Микробы заражают . (2003) 5: 387–95. DOI: 10.1016 / S1286-4579 (03) 00042-X
PubMed Аннотация | CrossRef Полный текст | Google Scholar
41. Ван Х, Го Х, Су Дж, Жуй Й, Чжэн В., Гао В. и др. Ингибирование Vpx-опосредованной деградации фактора транскрипции SAMHD1 и Vpr-хеликазы хозяина путем селективного нарушения сборки вирусной CRL4 (DCAF1) E3 убиквитинлигазы. Дж Вирол . (2017) 91: e00225-17 DOI: 10.1128 / JVI.00225-17
PubMed Аннотация | CrossRef Полный текст | Google Scholar
44. Wu Y, Zhou X, Barnes CO, DeLucia M, Cohen AE, Gronenborn AM, et al. Кристаллическая структура DDB1-DCAF1-Vpr-UNG2 показывает, как Vpr ВИЧ-1 направляет человеческий UNG2 к разрушению. Нат Струк Мол Биол . (2016) 23: 933–40. DOI: 10.1038 / nsmb.3284
PubMed Аннотация | CrossRef Полный текст | Google Scholar
45. Чжао Л.Дж., Мукерджи С., Нараян О.Биохимический механизм функции Vpr ВИЧ-I. Специфическое взаимодействие с клеточным белком. Дж. Биол. Хим. . (1994) 269: 15577–82.
PubMed Аннотация | Google Scholar
46. Грецка К., Хао Ц., Шун М. – Ц., Каур С., Свансон С. К., Флоренс Л. и др. ВИЧ-1 и ВИЧ-2 проявляют различные взаимодействия с белками репарации ДНК HLTF и UNG2. Труды Национальной академии наук . (2016) 113, E3921 – E30. DOI: 10.1073 / pnas.1605023113
PubMed Аннотация | CrossRef Полный текст | Google Scholar
47.Laguette N, Brégnard C, Hue P, Basbous J, Yatim A, Larroque M и др. Преждевременная активация комплекса SLX4 с помощью Vpr способствует остановке G2 / M и уходу от врожденного иммунного восприятия. Ячейка . (2014) 156: 134–45. DOI: 10.1016 / j.cell.2013.12.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
48. Lahouassa H, Blondot ML, Chauveau L, Chougui G, Morel M, Leduc M, et al. Vpr ВИЧ-1 разрушает ДНК-транслоказу HLTF в Т-клетках и макрофагах. Труды Национальной академии наук .(2016) 113: 5311–6. DOI: 10.1073 / pnas.1600485113
PubMed Аннотация | CrossRef Полный текст | Google Scholar
49. Янь Дж., Шун М.К., Хао С., Чжан Ю., Цянь Дж., Хрека К. и др. (2018). Vpr ВИЧ-1 перепрограммирует CLR4 DCAF1 E3 убиквитинлигазу для противодействия ограничению инфекции ВИЧ-1, опосредованной экзонуклеазой 1. мБио. 9 . DOI: 10.1128 / mBio.01732-18
PubMed Аннотация | CrossRef Полный текст | Google Scholar
50. Чжоу X, ДеЛусия М., Ан Дж.SLX4-SLX1, независимое от белка подавление белка MUS81-EME1 вирусным белком R (Vpr) ВИЧ-1. Журнал биологической химии . (2016) 291: 16936–47. DOI: 10.1074 / jbc.M116.721183
PubMed Аннотация | CrossRef Полный текст | Google Scholar
51. Rouzic EL, Belaïdouni N, Estrabaud E, Morel M, Rain J-C, Transy C., et al. (2007). Vpr ВИЧ1 останавливает клеточный цикл, рекрутируя DCAF1 / VprBP. рецептор убиквитинлигазы Cul4-DDB1. Cell Cycle , 6: 182–188.DOI: 10.4161 / cc.6.2.3732
PubMed Аннотация | CrossRef Полный текст | Google Scholar
52. Жако Дж., Ле Рузик Е., Майду-Пиндара П., Мэйзи М., Лефрер Дж. Дж., Данелуцци В. и др. (2009). Характеристика молекулярных детерминант первичных белков Vpr ВИЧ-1: влияние замен Q65R и R77Q на функции Vpr. PLOS ONE . 4: e7514. DOI: 10.1371 / journal.pone.0007514
PubMed Аннотация | CrossRef Полный текст | Google Scholar
53.Hansen EC, Ransom M, Hesselberth JR, Hosmane NN, Capoferri AA, Bruner KM, et al. Различная судьба урацилированной ДНК ВИЧ-1 при инфицировании клеток миелоидной линии. eLife . (2016) 5: e18447. DOI: 10.7554 / eLife.18447
PubMed Аннотация | CrossRef Полный текст | Google Scholar
54. Кеннеди Е.М., Даддача В., Слейтер Р., Гавеньяно С., Фроментин Е., Схинази Р.Ф. и др. Обильный неканонический dUTP, обнаруженный в первичных макрофагах человека, способствует его частому включению обратной транскриптазой ВИЧ-1. Дж. Биол. Хим. . (2011) 286: 25047–55. DOI: 10.1074 / jbc.M111.234047
PubMed Аннотация | CrossRef Полный текст | Google Scholar
55. Ланжевен К., Майду-Пиндара П., Аас П.А., Жако Г., Оттерлей М., Слуппхауг Г. и др. Vpr вируса иммунодефицита человека типа 1 модулирует клеточную экспрессию UNG2 посредством отрицательного транскрипционного эффекта. Дж Вирол . (2009) 83: 10256–63. DOI: 10.1128 / JVI.02654-08
PubMed Аннотация | CrossRef Полный текст | Google Scholar
56.Weil AF, Ghosh D, Zhou Y, Seiple L, McMahon MA, Spivak AM и др. Урацил-ДНК-гликозилаза инициирует деградацию кДНК ВИЧ-1, содержащей неправильно включенный dUTP, и предотвращает интеграцию вируса. Труды Национальной академии наук . (2013) 110: E448 – E57. DOI: 10.1073 / pnas.1219702110
PubMed Аннотация | CrossRef Полный текст | Google Scholar
57. Ан Дж., Ву Т., Новинце З., Герреро-Санторо Дж., Рапик-Отрин В., Гроненборн А.М. Vpr ВИЧ-1 загружает урацил ДНК-гликозилазу-2 на DCAF1, субъединицу распознавания субстрата убиквитинлигазы Cullin 4A-RING E3 для протеасомозависимой деградации. Журнал биологической химии . (2010) 285: 37333–41. DOI: 10.1074 / jbc.M110.133181
PubMed Аннотация | CrossRef Полный текст | Google Scholar
58. Бергер Г., Лоуренс М., Хью С., Нил С.Дж. (2014). Остановка клеточного цикла G2 / M коррелирует с взаимодействием лентивируса Vpr приматов с комплексом SLX4. Дж Вирол . 89: 230–240. DOI: 10.1128 / JVI.02307-14
PubMed Аннотация | CrossRef Полный текст | Google Scholar
59. Фрегосо О.И., Эмерман М.(2016). Активация ответа на повреждение ДНК является консервативной функцией Vpr ВИЧ-1 и ВИЧ-2, которая не зависит от рекрутирования SLX4. мБио.7 . DOI: 10.1128 / mBio.01433-16
PubMed Аннотация | CrossRef Полный текст | Google Scholar
60. Гринвуд Э.Д., Уильямсон Дж. С., Сенкевич А., Наамати А., Матесон, штат Нью-Джерси, Ленер П. Дж.. Беспорядочное нацеливание на клеточные белки с помощью Vpr управляет протеомным ремоделированием системного уровня при ВИЧ-1-инфекции. Cell Rep. (2019) 27: 1579–96e7.DOI: 10.1016 / j.celrep.2019.04.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
61. Achar YJ, Balogh D, Haracska L. Скоординированное ремоделирование белка и ДНК человеческим HLTF на остановленной репликационной вилке. Proc Natl Acad Sci USA . (2011) 108: 14073–8. DOI: 10.1073 / pnas.1101951108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
62. Кейзерс Г., Бакула Д., Петр М.А., Мадсен НГК, Теклу А., Мкртчян Г. и др. Регуляторные функции экзонуклеазы 1 человека (EXO1) в репликации ДНК с предполагаемой ролью при раке. Int J Mol Sci. (2018) 20:74. DOI: 10.3390 / ijms20010074
PubMed Аннотация | CrossRef Полный текст | Google Scholar
66. Мариджио Дж., Кох С., Чжан Дж., Вайднер-Глунде М., Рукерт Дж., Кати С. и др. Ядерный антиген, связанный с латентностью вируса герпеса саркомы Капоши (KSHV) (LANA), привлекает компоненты комплекса репарации MRN (Mre11-Rad50-NBS1) для модуляции сигнального пути врожденного иммунитета и вирусной латентности. PLoS Патог . (2017) 13: e1006335. DOI: 10,1371 / журнал.ппат.1006335
PubMed Аннотация | CrossRef Полный текст | Google Scholar
67. Тараканова В.Л., Леунг-Пинеда В., Хван С., Ян К.В., Мататалл К., Бассон М. и др. Гамма-герпесвирусная киназа активно инициирует ответ на повреждение ДНК, индуцируя фосфорилирование h3AX, чтобы способствовать репликации вируса. Клеточный микроб-хозяин . (2007) 1: 275–86. DOI: 10.1016 / j.chom.2007.05.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
68. Накай-Мураками К., Шимура М., Киномото М., Такидзава Ю., Токунага К., Тагучи Т. и др.Vpr ВИЧ-1 индуцирует ATM-зависимый клеточный сигнал с усиленной гомологичной рекомбинацией. Онкоген . (2007) 26: 477–86. DOI: 10.1038 / sj.onc.1209831
PubMed Аннотация | CrossRef Полный текст | Google Scholar
69. Wu ZH, Shi Y, Tibbetts RS, Miyamoto S. Молекулярная связь между киназой ATM и передачей сигналов NF-kappaB в ответ на генотоксические стимулы. Наука . (2006) 311: 1141–6. DOI: 10.1126 / science.1121513
PubMed Аннотация | CrossRef Полный текст | Google Scholar
70.Lv L, Wang Q, Xu Y, Tsao LC, Nakagawa T., Guo H, et al. Vpr нацелен на TET2 для деградации лигазой CRL4VprBP E3 для поддержания экспрессии IL-6 и усиления репликации ВИЧ-1. Мол Ячейки . (2018) 70: 961–70.e5. DOI: 10.1016 / j.molcel.2018.05.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
71. van Breugel PC, Robert EI, Mueller H, Decorsière A, Zoulim F, Hantz O, et al. Белок Х вируса гепатита В стимулирует экспрессию генов избирательно из внехромосомных ДНК-матриц. Гепатология . (2012) 56: 2116–24. DOI: 10.1002 / hep.25928
PubMed Аннотация | CrossRef Полный текст | Google Scholar
72. Maudet C, Sourisce A, Dragin L, Lahouassa H, Rain J-C, Bouaziz S, et al. Vpr ВИЧ-1 индуцирует деградацию ZIP и sZIP, адаптеров комплекса ремоделирования хроматина NuRD, путем захвата DCAF1 / VprBP. PLOS ONE . (2013) 8: e77320. DOI: 10.1371 / journal.pone.0077320
PubMed Аннотация | CrossRef Полный текст | Google Scholar
73.Епанчинцев А., Костанцо Ф., Раушендорф М.А., Капуто М., Йе Т, Доннио Л.М. и др. Белки А и В синдрома Кокейнса регулируют остановку транскрипции после генотоксического стресса, способствуя деградации ATF3. Мол Ячейки . (2017) 68: 1054–66.e6. DOI: 10.1016 / j.molcel.2017.11.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
74. Поли Г., Бресслер П., Кинтер А., Ду Е., Тиммер В. К., Рабсон А. и др. Интерлейкин 6 индуцирует экспрессию вируса иммунодефицита человека в инфицированных моноцитарных клетках в одиночку и в синергии с фактором некроза опухоли альфа за счет транскрипционных и посттранскрипционных механизмов. J Exp Med . (1990) 172: 151–8. DOI: 10.1084 / jem.172.1.151
PubMed Аннотация | CrossRef Полный текст | Google Scholar
75. Weissman D, Poli G, Fauci AS. Интерлейкин 10 блокирует репликацию ВИЧ в макрофагах путем ингибирования аутокринной петли фактора некроза опухоли α и индукции интерлейкина 6 вируса. Ретровирусы AIDS Res Hum . (1994) 10: 1199–206. DOI: 10.1089 / помощь.1994.10.1199
PubMed Аннотация | CrossRef Полный текст | Google Scholar
76.Чжан Ц., Чжао К., Шен Ц., Хань И, Гу И, Ли Х и др. Tet2 необходим для устранения воспаления путем привлечения Hdac2 для специфической репрессии IL-6. Природа . (2015) 525: 389–93. DOI: 10.1038 / природа15252
PubMed Аннотация | CrossRef Полный текст | Google Scholar
77. Чене Ид, Басюк Е., Лин Ю.Л., Трибуле Р., Кнезевич А., Шабл-Бессия С. и др. Suv39h2 и HP1γ ответственны за опосредованное хроматином подавление транскрипции ВИЧ-1 и латентность после интеграции. EMBO J .(2007) 26: 424–35. DOI: 10.1038 / sj.emboj.7601517
PubMed Аннотация | CrossRef Полный текст | Google Scholar
78. Часовникарова И.А., Тиммс Р.Т., Матесон Н.Дж., Уолс К., Антробус Р., Гёттгенс Б. и др. ГЕНОВАЯ ТИШИНА. Эпигенетическое молчание с помощью комплекса HUSH опосредует изменение положения в клетках человека. Наука. (2015) 348: 1481–5. DOI: 10.1126 / science.aaa7227
PubMed Аннотация | CrossRef Полный текст | Google Scholar
79. Chougui G, Munir-Matloob S, Matkovic R, Martin MM, Morel M, Lahouassa H, et al.Вирусный белок X ВИЧ-2 / SIV противодействует репрессорному комплексу HUSH. Нат Микробиол . (2018) 3: 891–7. DOI: 10.1038 / s41564-018-0179-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
80. Юрковецкий Л., Гуней М.Х., Ким К., Го С.Л., МакКоли С., Дофин А. и др. Белки вируса иммунодефицита приматов Vpx и Vpr противодействуют репрессии транскрипции провирусов комплексом HUSH. Нат Микробиол . (2018) 3: 1354. DOI: 10.1038 / s41564-018-0256-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
82.Хачия Р., Шиихаси Т., Сиракава И., Ивасаки Ю., Мацумура Ю., Оиси Ю. и др. Метилтрансфераза h4K9 Setdb1 регулирует TLR4-опосредованные воспалительные реакции в макрофагах. Научная репутация . (2016) 6: 28845. DOI: 10.1038 / srep28845
PubMed Аннотация | CrossRef Полный текст | Google Scholar
83. Xu G, Liu G, Xiong S, Liu H, Chen X, Zheng B. Гистонметилтрансфераза Smyd2 является негативным регулятором активации макрофагов, подавляя интерлейкин 6 (IL-6) и фактор некроза опухоли альфа (TNF-alpha ) производство. Дж. Биол. Хим. . (2015) 290: 5414–23. DOI: 10.1074 / jbc.M114.610345
PubMed Аннотация | CrossRef Полный текст | Google Scholar
84. Трибуле Р., Мари Б., Лин Ю.Л., Шабл-Бессия С., Беннассер И., Лебриганд К. и др. Подавление пути сайленсинга микроРНК ВИЧ-1 во время репликации вируса. Наука . (2007) 315: 1579–82. DOI: 10.1126 / science.1136319
PubMed Аннотация | CrossRef Полный текст | Google Scholar
86. Натанс Р., Чу С.Й., Серкина А.К., Лу С.К., Цао Х., Рана Т.М.Клеточная микроРНК и Р-тельца модулируют взаимодействия хозяин-ВИЧ-1. Мол Ячейки . (2009) 34: 696–709. DOI: 10.1016 / j.molcel.2009.06.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
88. Сили Дж. Дж., Бейкер Р. Г., Мохамед Дж., Брунс Т., Хайден М. С., Дешмук С. Д. и др. Индукция врожденной иммунной памяти через нацеливание микроРНК на факторы ремоделирования хроматина. Природа . (2018) 559: 114–9. DOI: 10.1038 / s41586-018-0253-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
89.Кейси Клоков Л., Шарифи Х. Дж., Вен X, Флэгг М., Фуруя А. К., Некорчук М. и др. Белок Vpr ВИЧ-1 нацелен на Dicer эндорибонуклеазы для протеасомной деградации, чтобы усилить инфекцию макрофагов. Вирусология . (2013) 444: 191–202. DOI: 10.1016 / j.virol.2013.06.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
91. Agosto LM, Uchil PD, Mothes W. Передача ВИЧ от клетки к клетке: влияние на патогенез и антиретровирусная терапия. Trends Microbiol .(2015) 23: 289–95. DOI: 10.1016 / j.tim.2015.02.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
92. Гуммулуру С., Пина Рамирес Н.Г., Акияма Х. CD169-зависимая клеточно-ассоциированная передача ВИЧ-1: фактор распространения вируса. J Infect Dis. (2014) 210 (Приложение 3): S641–7. DOI: 10.1093 / infdis / jiu442
PubMed Аннотация | CrossRef Полный текст | Google Scholar
93. Collins DR, Lubow J, Lukic Z, Mashiba M, Collins KL. Vpr способствует макрофагозависимой ВИЧ-1 инфекции CD4 + Т-лимфоцитов. PLOS Pathog . (2015) 11: e1005054. DOI: 10.1371 / journal.ppat.1005054
PubMed Аннотация | CrossRef Полный текст | Google Scholar
94. Машиба М, Коллинз Д.Р., Терри В.Х., Коллинз К.Л. Vpr преодолевает специфическое для макрофагов ограничение экспрессии Env ВИЧ-1 и продукции вирионов. Клеточный микроб-хозяин . (2014) 16: 722–35. DOI: 10.1016 / j.chom.2014.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
95. Muthumani K, Hwang DS, Choo AY, Mayilvahanan S, Dayes NS, Thieu KP, et al.ВИЧ-1 Vpr ингибирует созревание и активацию макрофагов и дендритных клеток in vitro. Инт Иммунол . (2005) 17: 103–16. DOI: 10.1093 / intimm / dxh290
PubMed Аннотация | CrossRef Полный текст | Google Scholar
96. Majumder B, Janket ML, Schafer EA, Schaubert K, Huang XL, Kan-Mitchell J, et al. Vpr вируса иммунодефицита человека 1 типа нарушает созревание дендритных клеток и активацию Т-клеток: последствия для ускользания от вирусного иммунитета. Дж Вирол . (2005) 79: 7990–8003.DOI: 10.1128 / JVI.79.13.7990-8003.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
97. Roux P, Alfieri C, Hrimech M, Cohen EA, Tanner JE. Активация факторов транскрипции NF-κB и NF-IL-6 белком R (Vpr) вируса иммунодефицита человека 1 типа индуцирует экспрессию интерлейкина-8. Дж Вирол . (2000) 74: 4658–65. DOI: 10.1128 / JVI.74.10.4658-4665.2000
PubMed Аннотация | CrossRef Полный текст | Google Scholar
98. Лю Р., Линь И, Цзя Р., Гэн Ю., Лян Ц., Тан Дж и др.Vpr ВИЧ-1 стимулирует передачу сигналов NF-kappaB и AP-1, активируя TAK1. Ретровирология . (2014) 11:45. DOI: 10.1186 / 1742-4690-11-45
PubMed Аннотация | CrossRef Полный текст | Google Scholar
101. Акияма Х., Миллер С.М., Эттингер С.Р., Белкина А.С., Снайдер-Каппионе Дж.Э., Гуммулуру С. Экспрессия РНК, содержащей интрон ВИЧ-1, вызывает активацию врожденного иммунитета и дисфункцию Т-клеток. Нац Коммуна . (2018) 9: 3450. DOI: 10.1038 / s41467-018-05899-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
104.Manganaro L, Hong P, Hernandez MM, Argyle D, Mulder LCF, Potla U и др. IL-15 регулирует восприимчивость CD4 + Т-клеток к ВИЧ-инфекции. Proc Natl Acad Sci USA . (2018) 115: E9659 – E67. DOI: 10.1073 / pnas.1806695115
PubMed Аннотация | CrossRef Полный текст | Google Scholar
105. Окумура А., Альсе Т., Лубьева Б., Эзель Х., Штребель К., Питха П.М.. Вспомогательные белки ВИЧ-1 VPR и Vif модулируют противовирусный ответ, нацеливая IRF-3 на деградацию. Вирусология .(2008) 373: 85–97. DOI: 10.1016 / j.virol.2007.10.042
PubMed Аннотация | CrossRef Полный текст | Google Scholar
106. Харман А.Н., Наср Н., Фитхам А., Галоян А., Альшехри А.А., Рамбуквелле Д. и др. ВИЧ блокирует индукцию интерферона в дендритных клетках и макрофагах человека за счет нарушения регуляции TBK1. Дж Вирол . (2015) 89: 6575–84. DOI: 10.1128 / JVI.00889-15
PubMed Аннотация | CrossRef Полный текст | Google Scholar
107. Акияма Х., Рамирес Н.Г., Гудхети М.В., Гуммулуру С.CD169-опосредованный перенос ВИЧ в инвагинации плазматической мембраны в дендритных клетках ослабляет эффективность широко нейтрализующих антител против gp120. PLoS Патог . (2015) 11: e1004751. DOI: 10.1371 / journal.ppat.1004751
PubMed Аннотация | CrossRef Полный текст | Google Scholar
108. Puryear WB, Akiyama H, Geer SD, Ramirez NP, Yu X, Reinhard BM, et al. Интерферон-индуцируемый механизм опосредованного дендритными клетками распространения ВИЧ-1 зависит от Siglec-1 / CD169. PLoS Патог . (2013) 9: e1003291. DOI: 10.1371 / journal.ppat.1003291
PubMed Аннотация | CrossRef Полный текст | Google Scholar
109. Криббс С.К., Леннокс Дж., Калиендо А.М., Браун Л.А., Гуидо Д.М. Здоровые ВИЧ-1-инфицированные люди, получающие высокоактивную антиретровирусную терапию, содержат ВИЧ-1 в своих альвеолярных макрофагах. Ретровирусы AIDS Res Hum . (2015) 31: 64–70. DOI: 10.1089 / aid.2014.0133
PubMed Аннотация | CrossRef Полный текст | Google Scholar
110.Эстес Дж. Д., Китио С., Ссали Ф., Свейнсон Л., Макамдоп К. Н., Дель Прете Г. К. и др. Определение бремени вируса СПИДа для всего организма с последствиями для лечебных стратегий. Нат Мед . (2017) 23: 1271–6. DOI: 10,1038 / нм 4411
PubMed Аннотация | CrossRef Полный текст | Google Scholar
111. Honeycutt JB, Thayer WO, Baker CE, Ribeiro RM, Lada SM, Cao Y, et al. Персистенция ВИЧ в тканевых макрофагах гуманизированных мышей, содержащих только миелоиды, во время антиретровирусной терапии. Нат Мед . (2017) 23: 638–43.DOI: 10,1038 / нм 4319
PubMed Аннотация | CrossRef Полный текст | Google Scholar
112. Ламерс С.Л., Роуз Р., Майджи Э., Агсалда-Гарсия М., Нолан Д.Д., Фогель Г.Б. и др. ДНК ВИЧ часто присутствует в патологических тканях, оцениваемых при вскрытии, у пациентов, получавших комбинированную антиретровирусную терапию, с неопределяемой вирусной нагрузкой. Дж Вирол . (2016) 90: 8968–83. DOI: 10.1128 / JVI.00674-16
PubMed Аннотация | CrossRef Полный текст | Google Scholar
113.McElrath MJ, Smythe K, Randolph-Habecker J, Melton KR, Goodpaster TA, Hughes SM и др. Всесторонняя оценка клеток-мишеней ВИЧ в дистальном отделе кишечника человека предполагает повышение восприимчивости к ВИЧ в области заднего прохода. J Синдр иммунодефицита Acquir . (2013) 63: 263–71. DOI: 10.1097 / QAI.0b013e3182898392
PubMed Аннотация | CrossRef Полный текст | Google Scholar
114. Юкл С.А., Синклер Э., Сомсук М., Хант П.В., Эплинг Л., Киллиан М. и др. (2014). Сравнение методов измерения ректального уровня ВИЧ позволяет предположить, что ДНК ВИЧ находится в клетках, отличных от CD4 + Т-клеток, включая миелоидные клетки. СПИД . 28: 439–442. DOI: 10.1097 / QAD.0000000000000166
PubMed Аннотация | CrossRef Полный текст | Google Scholar
115. Арайнга М., Эдагва Б., Мосли Р.Л., Полуэктова Л.Ю., Горантла С., Гендельман Х.Э. Зрелый макрофаг является основным клеточным резервуаром ВИЧ-1 у гуманизированных мышей после лечения антиретровирусной терапией длительного действия. Ретровирология . (2017) 14:17. DOI: 10.1186 / s12977-017-0344-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Белок Vpr из ВИЧ-1: различные роли в жизненном цикле вируса | Ретровирология
Tristem M, Marshall C, Karpas A, Hill F: Эволюция лентивирусов приматов: данные vpx и vpr. Embo J. 1992, 11: 3405-3412.
PubMed Central CAS PubMed Google Scholar
Hoch J, Lang SM, Weeger M, Stahl-Hennig C, Coulibaly C, Dittmer U, Hunsmann G, Fuchs D, Muller J, Sopper S и др.: Мутант с делецией vpr вируса иммунодефицита обезьян вызывает СПИД у макак-резусов. J Virol. 1995, 69: 4807-4813.
PubMed Central CAS PubMed Google Scholar
Lang SM, Weeger M, Stahl-Hennig C, Coulibaly C, Hunsmann G, Muller J, Muller-Hermelink H, Fuchs D, Wachter H, Daniel MM, et al: Важность vpr для инфицирования резуса обезьяны с вирусом обезьяньего иммунодефицита. J Virol. 1993, 67: 902-912.
PubMed Central CAS PubMed Google Scholar
Гиббс Дж.С., Лакнер А.А., Ланг С.М., Саймон М.А., Сегал П.К., Дэниел М.Д., Десрозье Р.С.: прогрессирование до СПИДа в отсутствие гена для vpr или vpx. J Virol. 1995, 69: 2378-2383.
PubMed Central CAS PubMed Google Scholar
Hirsch VM, Sharkey ME, Brown CR, Brichacek B, Goldstein S, Wakefield J, Byrum R, Elkins WR, Hahn BH, Lifson JD, Stevenson M: Vpx требуется для распространения и патогенеза ВИО (SM ) PBj: свидетельство макрофагозависимой вирусной амплификации.Nat Med. 1998, 4: 1401-1408. 10.1038 / 3992.
CAS PubMed Google Scholar
Henklein P, Bruns K, Sherman MP, Tessmer U, Licha K, Kopp J, de Noronha CM, Greene WC, Wray V, Schubert U: Функциональная и структурная характеристика синтетического Vpr ВИЧ-1, который трансдуцирует клетки , локализуется в ядре и вызывает остановку клеточного цикла G2. J Biol Chem. 2000, 275: 32016-32026. 10.1074 / jbc.M004044200.
CAS PubMed Google Scholar
Schuler W, Wecker K, de Rocquigny H, Baudat Y, Sire J, Roques BP: ЯМР-структура (52–96) C-концевого домена (52–96) C-концевого домена регуляторного белка ВИЧ-1 Vpr: молекулярное понимание его биологических функций. J Mol Biol. 1999, 285: 2105-2117. 10.1006 / jmbi.1998.2381.
CAS PubMed Google Scholar
Wecker K, Roques BP: ЯМР-структура (1–51) N-концевого домена регуляторного белка ВИЧ-1 Vpr. Eur J Biochem. 1999, 266: 359-369.10.1046 / j.1432-1327.1999.00858.x.
CAS PubMed Google Scholar
Wecker K, Morellet N, Bouaziz S, Roques BP: Структура ЯМР регуляторного белка ВИЧ-1 Vpr в h3O / трифторэтаноле. Сравнение с N-концевым (1–51) и C-концевым (52–96) доменами Vpr. Eur J Biochem. 2002, 269: 3779-3788. 10.1046 / j.1432-1033.2002.03067.x.
CAS PubMed Google Scholar
Яо XJ, Subbramanian RA, Rougeau N, Boisvert F, Bergeron D, Cohen EA: Мутагенный анализ Vpr вируса иммунодефицита человека типа 1: роль предсказанной N-концевой альфа-спиральной структуры в ядерной локализации Vpr и включении вирионов. J Virol. 1995, 69: 7032-7044.
PubMed Central CAS PubMed Google Scholar
Bourbigot S, Beltz H, Denis J, Morellet N, Roques BP, Mely Y, Bouaziz S: C-концевой домен VPR принимает антипараллельную димерную структуру в растворе через свой домен, подобный лейциновой молнии.Biochem J. 2004,
Google Scholar
Ван Л., Мукерджи С., Нараян О., Чжао Л.Дж.: Характеристика домена, подобного лейциновой молнии, в белке Vpr вируса иммунодефицита человека типа 1. Ген. 1996, 178: 7-13. 10.1016 / 0378-1119 (96) 00312-5.
CAS PubMed Google Scholar
Mahalingam S, Ayyavoo V, Patel M, Kieber-Emmons T, Weiner DB: Ядерный импорт, включение вирионов и остановка / дифференцировка клеточного цикла опосредуются отдельными функциональными доменами Vpr вируса иммунодефицита человека 1 типа.J Virol. 1997, 71: 6339-6347.
PubMed Central CAS PubMed Google Scholar
Чжао Л.Дж., Ван Л., Мукерджи С., Нараян О.: Биохимический механизм функции Vpr ВИЧ-1. Олигомеризация опосредована N-концевым доменом. J Biol Chem. 1994, 269: 32131-32137.
CAS PubMed Google Scholar
Морелле Н., Буазиз С., Петижан П., Рокес Б. П.: ЯМР-структура регуляторного белка ВИЧ-1 VPR.J Mol Biol. 2003, 327: 215-227. 10.1016 / S0022-2836 (03) 00060-3.
CAS PubMed Google Scholar
Брунс К., Фоссен Т., Рэй В., Хенклейн П., Тессмер Ю., Шуберт Ю.: Структурная характеристика N-конца Vpr ВИЧ-1: свидетельство цис / транс-пролиновой изомерии. J Biol Chem. 2003, 278: 43188-43201. 10.1074 / jbc.M305413200.
CAS PubMed Google Scholar
Zander K, Sherman MP, Tessmer U, Bruns K, Wray V, Prechtel AT, Schubert E, Henklein P, Luban J, Neidleman J и др .: Циклофилин A взаимодействует с Vpr ВИЧ-1 и необходим для его функциональной экспрессии. J Biol Chem. 2003, 278: 43202-43213. 10.1074 / jbc.M305414200.
CAS PubMed Google Scholar
Шерман М.П., Шуберт У., Уильямс С.А., де Норонья С.М., Крейсберг Дж.Ф., Хенклейн П., Грин В.К.: ВИЧ-1 Vpr проявляет естественные белковые трансдуцирующие свойства: последствия для вирусного патогенеза.Вирусология. 2002, 302: 95-105. 10.1006 / viro.2002.1576.
CAS PubMed Google Scholar
Kichler A, Pages JC, Leborgne C, Druillennec S, Lenoir C, Coulaud D, Delain E, Le Cam E, Roques BP, Danos O: эффективная трансфекция ДНК, опосредованная C-концевым доменом иммунодефицита человека вирусный вирусный белок 1 типа R. J Virol. 2000, 74: 5424-5431. 10.1128 / JVI.74.12.5424-5431.2000.
PubMed Central CAS PubMed Google Scholar
Coeytaux E, Coulaud D, Le Cam E, Danos O, Kichler A: Катионная амфипатическая альфа-спираль вирусного белка R (Vpr) ВИЧ-1 связывается с нуклеиновыми кислотами, проницаемо для мембран и эффективно трансфицирует клетки. J Biol Chem. 2003, 278: 18110-18116. 10.1074 / jbc.M300248200.
CAS PubMed Google Scholar
Аккола М.А., Буковский А.А., Джонс М.С., Готтлингер Х.Г.: Консервативный дилейцин-содержащий мотив в p6 (gag) регулирует ассоциацию частиц Vpx и Vpr вирусов иммунодефицита обезьян SIV (mac) и SIV (agm) .J Virol. 1999, 73: 9992-9999.
PubMed Central CAS PubMed Google Scholar
Bachand F, Yao XJ, Hrimech M, Rougeau N, Cohen EA: Включение Vpr в вирус иммунодефицита человека типа 1 требует прямого взаимодействия с доменом p6 предшественника p55 gag. J Biol Chem. 1999, 274: 9083-9091. 10.1074 / jbc.274.13.9083.
CAS PubMed Google Scholar
Jenkins Y, Sanchez PV, Meyer BE, Malim MH: Ядерный экспорт вируса иммунодефицита человека типа 1 Vpr не требуется для упаковки вирионов. J Virol. 2001, 75: 8348-8352. 10.1128 / JVI.75.17.8348-8352.2001.
PubMed Central CAS PubMed Google Scholar
Selig L, Pages JC, Tanchou V, Preveral S, Berlioz-Torrent C, Liu LX, Erdtmann L, Darlix J, Benarous R, Benichou S: Взаимодействие с доменом p6 предшественника gag опосредует включение в вирионы белков Vpr и Vpx лентивирусов приматов.J Virol. 1999, 73: 592-600.
PubMed Central CAS PubMed Google Scholar
Singh SP, Tomkowicz B, Lai D, Cartas M, Mahalingam S, Kalyanaraman VS, Murali R, Srinivasan A: Функциональная роль остатков, соответствующих спиральному домену II (аминокислоты от 35 до 46) вируса иммунодефицита человека тип 1 Впр. J Virol. 2000, 74: 10650-10657. 10.1128 / JVI.74.22.10650-10657.2000.
PubMed Central CAS PubMed Google Scholar
Kondo E, Gottlinger HG: консервативная последовательность LXXLF является основной детерминантой в p6gag, необходимой для включения Vpr вируса иммунодефицита человека типа 1. J Virol. 1996, 70: 159-164.
PubMed Central CAS PubMed Google Scholar
Accola MA, Ohagen A, Gottlinger HG: Выделение ядер вируса иммунодефицита человека типа 1: сохранение vpr в отсутствие p6 (gag) [ссылка в процессе]. J Virol. 2000, 74: 6198-6202.10.1128 / JVI.74.13.6198-6202.2000.
PubMed Central CAS PubMed Google Scholar
Велкер Р., Хоэнберг Х., Тессмер У., Хукхагель С., Краусслих Х.Г.: Биохимический и структурный анализ изолированных зрелых ядер вируса иммунодефицита человека типа 1. J Virol. 2000, 74: 1168-1177. 10.1128 / JVI.74.3.1168-1177.2000.
PubMed Central CAS PubMed Google Scholar
Zhang S, Pointer D, Singer G, Feng Y, Park K, Zhao LJ: Прямое связывание с нуклеиновыми кислотами с помощью Vpr вируса иммунодефицита человека типа 1. Ген. 1998, 212: 157-166. 10.1016 / S0378-1119 (98) 00178-4.
CAS PubMed Google Scholar
de Rocquigny H, Caneparo A, Delaunay T., Bischerour J, Mouscadet JF, Roques BP: Взаимодействия C-конца вирусного белка R с нуклеиновыми кислотами модулируются его N-концом. Eur J Biochem.2000, 267: 3654-3660. 10.1046 / j.1432-1327.2000.01397.x.
CAS PubMed Google Scholar
de Rocquigny H, Petitjean P, Tanchou V, Decimo D, Drouot L, Delaunay T., Darlix JL, Roques BP: цинковые пальцы нуклеокапсидного белка NCp7 ВИЧ напрямую взаимодействуют с вирусным регуляторным белком Vpr. J Biol Chem. 1997, 272: 30753-30759. 10.1074 / jbc.272.49.30753.
CAS PubMed Google Scholar
Sato A, Yoshimoto J, Isaka Y, Miki S, Suyama A, Adachi A, Hayami M, Fujiwara T., Yoshie O: Доказательства прямой связи Vpr и матричного белка p17 в вирионе ВИЧ-1. Вирусология. 1996, 220: 208-212. 10.1006 / viro.1996.0302.
CAS PubMed Google Scholar
Muller B, Tessmer U, Schubert U, Krausslich HG: Белок Vpr вируса иммунодефицита человека типа 1 включается в вирион в значительно меньших количествах, чем gag, и фосфорилируется в инфицированных клетках.J Virol. 2000, 74: 9727-9731. 10.1128 / JVI.74.20.9727-9731.2000.
PubMed Central CAS PubMed Google Scholar
Ву Х, Лю Х., Сяо Х., Ким Дж., Сешайя П., Нацулис Дж., Боке Дж. Д., Хан Б. Х., Каппес Дж. К. Нацеливание чужеродных белков на частицы вируса иммунодефицита человека посредством слияния с Vpr и Vpx. J Virol. 1995, 69: 3389-3398.
PubMed Central CAS PubMed Google Scholar
Яо XJ, Kobinger G, Dandache S, Rougeau N, Cohen E: гибридные белки Vpr-хлорамфениколацетилтрансферазы ВИЧ-1: требования к последовательности для включения вириона и анализ противовирусного эффекта. Gene Ther. 1999, 6: 1590-1599. 10.1038 / sj.gt.3300988.
CAS PubMed Google Scholar
Wu X, Liu H, Xiao H, Conway JA, Hunter E, Kappes JC: функциональные RT и IN, включенные в частицы ВИЧ-1 независимо от белка-предшественника Gag / Pol.Embo J. 1997, 16: 5113-5122. 10.1093 / emboj / 16.16.5113.
PubMed Central CAS PubMed Google Scholar
Лю Х., Ву Х, Сяо Х., Каппес Дж. К.: Нацеливание на белок интегразы вируса иммунодефицита человека (ВИЧ) типа 2 в ВИЧ типа 1. J Virol. 1999, 73: 8831-8836.
PubMed Central CAS PubMed Google Scholar
Wu X, Liu H, Xiao H, Conway JA, Hehl E, Kalpana GV, Prasad V, Kappes JC: Белок интегразы вируса иммунодефицита человека типа 1 способствует обратной транскрипции посредством специфических взаимодействий с комплексом обратной транскрипции нуклеопротеинов.J Virol. 1999, 73: 2126-2135.
PubMed Central CAS PubMed Google Scholar
Padow M, Lai L, Deivanayagam C, DeLucas LJ, Weiss RB, Dunn DM, Wu X, Kappes JC: Репликация химерного вируса иммунодефицита человека типа 1 (ВИЧ-1), содержащего интегразу ВИЧ-2 (IN ): естественно выбранные мутации в IN усиливают синтез ДНК. J Virol. 2003, 77: 11050-11059. 10.1128 / JVI.77.20.11050-11059.2003.
PubMed Central CAS PubMed Google Scholar
McDonald D, Vodicka MA, Lucero G, Svitkina TM, Borisy GG, Emerman M, Hope TJ: Визуализация внутриклеточного поведения ВИЧ в живых клетках. J Cell Biol. 2002, 159: 441-452. 10.1083 / jcb.200203150.
PubMed Central CAS PubMed Google Scholar
McDonald D, Wu L, Bohks SM, KewalRamani VN, Unutmaz D, Hope TJ: Рекрутирование ВИЧ и его рецепторов на соединения дендритных клеток с Т-клетками. Наука. 2003, 300: 1295-1297.10.1126 / science.1084238.
CAS PubMed Google Scholar
Farnet CM, Haseltine WA: Определение вирусных белков, присутствующих в преинтеграционном комплексе вируса иммунодефицита человека 1 типа. J Virol. 1991, 65: 1910-1915.
PubMed Central CAS PubMed Google Scholar
Fassati A, Goff SP: Характеристика внутриклеточных комплексов обратной транскрипции вируса иммунодефицита человека типа 1.J Virol. 2001, 75: 3626-3635. 10.1128 / JVI.75.8.3626-3635.2001.
PubMed Central CAS PubMed Google Scholar
Miller MD, Farnet CM, Bushman FD: Преинтеграционные комплексы вируса иммунодефицита человека типа 1: исследования организации и состава. J Virol. 1997, 71: 5382-5390.
PubMed Central CAS PubMed Google Scholar
Нермут М.В., Фассати А: Структурные анализы очищенных внутриклеточных комплексов обратной транскрипции вируса иммунодефицита человека типа 1.J Virol. 2003, 77: 8196-8206. 10.1128 / JVI.77.15.8196-8206.2003.
PubMed Central CAS PubMed Google Scholar
Букринский М.И., Хаггерти С., Демпси М.П., Шарова Н., Аджубель А., Шпиц Л., Льюис П., Гольдфарб Д., Эмерман М., Стивенсон М. Сигнал ядерной локализации в матричном белке ВИЧ-1, который управляет инфекцией неделящиеся клетки [см. комментарии]. Природа. 1993, 365: 666-669. 10.1038 / 365666a0.
CAS PubMed Google Scholar
Goff SP: Внутриклеточный трафик ретровирусных геномов на ранней стадии инфекции: вирусное использование клеточных путей. J Gene Med. 2001, 3: 517-528. 10.1002 / 1521-2254 (200111) 3: 6 <517 :: AID-JGM234> 3.0.CO; 2-E.
CAS PubMed Google Scholar
Zhang H, Dornadula G, Orenstein J, Pomerantz RJ: Морфологические изменения вирионов вируса иммунодефицита человека типа 1, вторичные по отношению к обратной транскрипции внутривириона: данные, указывающие на то, что обратная транскрипция может не происходить в интактном вирусном ядре.J Hum Virol. 2000, 3: 165-172.
CAS PubMed Google Scholar
Fassati A, Gorlich D, Harrison I., Zaytseva L, Mingot JM: Ядерный импорт внутриклеточных комплексов обратной транскрипции ВИЧ-1 опосредуется импортином 7. Embo J. 2003, 22: 3675-3685. 10.1093 / emboj / cdg357.
PubMed Central CAS PubMed Google Scholar
Старк Л.А., Хей RT: Вирусный белок R (Vpr) вируса иммунодефицита человека типа 1 (ВИЧ-1) взаимодействует с Lys-тРНК-синтетазой: последствия для примирования обратной транскрипции ВИЧ-1.J Virol. 1998, 72: 3037-3044.
PubMed Central CAS PubMed Google Scholar
Mansky LM, Temin HM: Более низкая частота мутаций in vivo вируса иммунодефицита человека типа 1, чем предсказанная на основе точности очищенной обратной транскриптазы. J Virol. 1995, 69: 5087-5094.
PubMed Central CAS PubMed Google Scholar
Мански Л.М.: На частоту мутаций вируса иммунодефицита человека типа 1 влияет ген vpr.Вирусология. 1996, 222: 391-400. 10.1006 / viro.1996.0436.
CAS PubMed Google Scholar
Chen R, Le Rouzic E, Kearney JA, Mansky LM, Benichou S: Vpr-опосредованное включение UNG2 в частицы ВИЧ-1 необходимо для модуляции скорости мутации вируса и для репликации в макрофагах. J Biol Chem. 2004, 279: 28419-28425. 10.1074 / jbc.M403875200.
CAS PubMed Google Scholar
Mansky LM, Preveral S, Selig L, Benarous R, Benichou S: Взаимодействие Vpr с урацил-ДНК-гликозилазой модулирует скорость мутаций вируса иммунодефицита человека 1 типа in vivo и мутаций. J Virol. 2000, 74: 7039-7047. 10.1128 / JVI.74.15.7039-7047.2000.
PubMed Central CAS PubMed Google Scholar
Agostini I, Navarro JM, Bouhamdan M, Willetts K, Rey F, Spire B, Vigne R, Pomerantz R, Sire J: Коактиватор Vpr ВИЧ-1 вызывает конформационные изменения в TFIIB.FEBS Lett. 1999, 450: 235-239. 10.1016 / S0014-5793 (99) 00501-3.
CAS PubMed Google Scholar
Jacotot E, Ravagnan L, Loeffler M, Ferri KF, Vieira HL, Zamzami N, Costantini P, Druillennec S, Hoebeke J, Briand JP и др.: Вирусный белок R ВИЧ-1 индуцирует апоптоз через прямое влияние на проницаемость митохондрий переходной поры. J Exp Med. 2000, 191: 33-46. 10.1084 / jem.191.1.33.
PubMed Central CAS PubMed Google Scholar
Klarmann GJ, Chen X, North TW, Preston BD: Включение урацила в минус-цепь ДНК влияет на специфичность инициации синтеза плюсовой цепи во время обратной транскрипции лентивируса. J Biol Chem. 2003, 278: 7902-7909. 10.1074 / jbc.M207223200.
CAS PubMed Google Scholar
Чен Р., Ван Х., Мански Л. М.: Роль урацил-ДНК-гликозилазы и дУТФазы в репликации вируса. J Gen Virol. 2002, 83: 2339-2345.
CAS PubMed Google Scholar
Sheehy AM, Gaddis NC, Choi JD, Malim MH: Выделение человеческого гена, который подавляет инфекцию ВИЧ-1 и подавляется вирусным белком Vif. Природа. 2002, 418: 646-650. 10.1038 / природа00939.
CAS PubMed Google Scholar
Zhang H, Yang B, Pomerantz RJ, Zhang C, Arunachalam SC, Gao L: Цитидиндезаминаза CEM15 индуцирует гипермутацию во вновь синтезированной ДНК ВИЧ-1. Природа. 2003, 424: 94-98. 10.1038 / природа01707.
PubMed Central CAS PubMed Google Scholar
Мариани Р., Чен Д., Шрофельбауэр Б., Наварро Ф., Кониг Р., Боллман Б., Мунк С., Нимарк-МакМахон Н., Ландау Н. Р.: Специфичное для видов исключение APOBEC3G из вирионов ВИЧ-1 с помощью Vif. Клетка. 2003, 114: 21-31. 10.1016 / S0092-8674 (03) 00515-4.
CAS PubMed Google Scholar
Mangeat B, Turelli P, Caron G, Friedli M, Perrin L, Trono D: Широкая антиретровирусная защита APOBEC3G человека посредством летального редактирования возникающих обратных транскриптов.Природа. 2003, 424: 99-103. 10.1038 / природа01709.
CAS PubMed Google Scholar
Lecossier D, Bouchonnet F, Clavel F, Hance AJ: Гипермутация ДНК ВИЧ-1 в отсутствие белка Vif. Наука. 2003, 300: 1112-10.1126 / science.1083338.
CAS PubMed Google Scholar
Коэн О.Дж., Фаучи А.С.: Современные стратегии лечения ВИЧ-инфекции.Adv Intern Med. 2001, 46: 207-246.
CAS PubMed Google Scholar
Eckstein DA, Sherman MP, Penn ML, Chin PS, De Noronha CM, Greene WC, Goldsmith MA: ВИЧ-1 Vpr увеличивает вирусную нагрузку, способствуя инфицированию тканевых макрофагов, но не неделящим CD4 + Т-клеткам. J Exp Med. 2001, 194: 1407-1419. 10.1084 / jem.194.10.1407.
PubMed Central CAS PubMed Google Scholar
Гребер У. Ф., Фассати А: Ядерный импорт вирусных ДНК геномов. Движение. 2003, 4: 136-143.
CAS PubMed Google Scholar
Коннор Р.И., Чен Б.К., Чоу С., Ландау Н.Р.: Vpr требуется для эффективной репликации вируса иммунодефицита человека типа 1 в мононуклеарных фагоцитах. Вирусология. 1995, 206: 935-944. 10.1006 / viro.1995.1016.
CAS PubMed Google Scholar
Gallay P, Stitt V, Mundy C, Oettinger M, Trono D: Роль кариоферинового пути в ядерном импорте вируса иммунодефицита человека 1 типа. J Virol. 1996, 70: 1027-1032.
PubMed Central CAS PubMed Google Scholar
Heinzinger NK, Bukinsky MI, Haggerty SA, Ragland AM, Kewalramani V, Lee MA, Gendelman HE, Ratner L, Stevenson M, Emerman M: Белок Vpr вируса иммунодефицита человека 1 типа влияет на ядерную локализацию вируса. нуклеиновые кислоты в неделящихся клетках-хозяевах.Proc Natl Acad Sci U S. A. 1994, 91: 7311-7315.
PubMed Central CAS PubMed Google Scholar
Le Rouzic E, Mousnier A, Rustum C, Stutz F, Hallberg E, Dargemont C, Benichou S. Присоединение Vpr ВИЧ-1 к ядерной оболочке опосредуется взаимодействием с нуклеопорином hCG1. J Biol Chem. 2002, 277: 45091-45098. 10.1074 / jbc.M207439200.
CAS PubMed Google Scholar
Попов С., Рексач М., Ратнер Л., Блобель Г., Букринский М.: Вирусный белок R регулирует стыковку прединтеграционного комплекса ВИЧ-1 с комплексом ядерной поры. J Biol Chem. 1998, 273: 13347-13352. 10.1074 / jbc.273.21.13347.
CAS PubMed Google Scholar
Popov S, Rexach M, Zybarth G, Reiling N, Lee MA, Ratner L, Lane CM, Moore MS, Blobel G, Bukrinsky M: вирусный белок R регулирует ядерный импорт пре-интеграции ВИЧ-1 сложный.Embo J. 1998, 17: 909-917. 10.1093 / emboj / 17.4.909.
PubMed Central CAS PubMed Google Scholar
Vodicka MA, Koepp DM, Silver PA, Emerman M: ВИЧ-1 Vpr взаимодействует с ядерным транспортным путем, способствуя заражению макрофагами. Genes Dev. 1998, 12: 175-185.
PubMed Central CAS PubMed Google Scholar
Fouchier RA, Meyer BE, Simon JH, Fischer U, Albright AV, Gonzalez-Scarano F, Malim MH: Взаимодействие белка Vpr вируса иммунодефицита человека типа 1 с комплексом ядерных пор.J Virol. 1998, 72: 6004-6013.
PubMed Central CAS PubMed Google Scholar
Fouchier RA, Malim MH: Ядерный импорт преинтеграционных комплексов вируса иммунодефицита человека типа 1. Adv Virus Res. 1999, 52: 275-299.
CAS PubMed Google Scholar
Каллен BR: Путешествие в центр камеры. Клетка. 2001, 105: 697-700. 10.1016 / S0092-8674 (01) 00392-0.
CAS PubMed Google Scholar
Букринский М., Аджубей А: Вирусный белок R ВИЧ-1. Rev Med Virol. 1999, 9: 39-49. 10.1002 / (SICI) 1099-1654 (199901/03) 9: 1 <39 :: AID-RMV235> 3.3.CO; 2-V.
CAS PubMed Google Scholar
Букринская А., Бричачек Б., Манн А., Стивенсон М.: Создание функционального комплекса обратной транскрипции вируса иммунодефицита человека типа 1 (ВИЧ-1) включает цитоскелет.J Exp Med. 1998, 188: 2113-2125. 10.1084 / jem.188.11.2113.
PubMed Central CAS PubMed Google Scholar
Waldhuber MG, Bateson M, Tan J, Greenway AL, McPhee DA: Исследования с использованием слитых белков GFP-Vpr: индукция апоптоза, но устранение остановки клеточного цикла, несмотря на ядерную мембрану или ядерную локализацию. Вирусология. 2003, 313: 91-104. 10.1016 / S0042-6822 (03) 00258-7.
CAS PubMed Google Scholar
Kamata M, Aida Y: Два предполагаемых альфа-спиральных домена Vpr вируса иммунодефицита человека типа 1 опосредуют ядерную локализацию по крайней мере двумя механизмами. J Virol. 2000, 74: 7179-7186. 10.1128 / JVI.74.15.7179-7186.2000.
PubMed Central CAS PubMed Google Scholar
Depienne C, Roques P, Creminon C, Fritsch L, Casseron R, Dormont D, Dargemont C, Benichou S: Распределение в клетках и кариофильные свойства белков матрицы, интегразы и Vpr из вирусов иммунодефицита человека и обезьян .Exp Cell Res. 2000, 260: 387-395. 10.1006 / excr.2000.5016.
CAS PubMed Google Scholar
Cronshaw JM, Krutchinsky AN, Zhang W, Chait BT, Matunis MJ: Протеомный анализ ядерного порового комплекса млекопитающих. J Cell Biol. 2002, 26: 915-927. 10.1083 / jcb.200206106.
Google Scholar
Rout MP, Aitchison JD: Ядерный поровый комплекс как транспортная машина.J Biol Chem. 2001, 276: 16593-16596. 10.1074 / jbc.R100015200.
CAS PubMed Google Scholar
Рейл Х., Буковский А.А., Гелдерблом Х.Р., Готтлингер Х.Г.: Эффективная репликация ВИЧ-1 может происходить в отсутствие вирусного матричного белка. Embo J. 1998, 17: 2699-2708. 10.1093 / emboj / 17.9.2699.
PubMed Central CAS PubMed Google Scholar
Хаффар О.К., Попов С., Дубровский Л., Агостини И., Танг Х., Пушкарский Т., Надлер С.Г., Букринский М. Два сигнала ядерной локализации в матричном белке ВИЧ-1 регулируют ядерный импорт предшественника ВИЧ-1. -интеграционный комплекс.J Mol Biol. 2000, 299: 359-368. 10.1006 / jmbi.2000.3768.
CAS PubMed Google Scholar
Dupont S, Sharova N, DeHoratius C, Virbasius CM, Zhu X, Bukrinskaya AG, Stevenson M, Green MR: Новая ядерная экспортная активность в матричном белке ВИЧ-1, необходимая для репликации вируса. Природа. 1999, 402: 681-685. 10.1038 / 45272.
CAS PubMed Google Scholar
de Noronha CM, Sherman MP, Lin HW, Cavrois MV, Moir RD, Goldman RD, Greene WC: динамические нарушения в архитектуре и целостности ядерной оболочки, вызванные Vpr ВИЧ-1. Наука. 2001, 294: 1105-1108. 10.1126 / science.1063957.
CAS PubMed Google Scholar
Сегура-Тоттен М, Уилсон К.Л .: Вирусология. ВИЧ — нарушение правил проникновения в ядерную среду. Наука. 2001, 294: 1016-1017. 10.1126 / science.1066729.
CAS PubMed Google Scholar
Лу ИЛ, Спирмен П., Ратнер Л.: Локализация вирусного белка R вируса иммунодефицита человека типа 1 в инфицированных клетках и вирионах. J Virol. 1993, 67: 6542-6550.
PubMed Central CAS PubMed Google Scholar
Zhou Y, Lu Y, Ratner L: Остатки аргинина на С-конце Vpr ВИЧ-1 важны для ядерной локализации и остановки клеточного цикла. Вирусология. 1998, 242: 414-424. 10.1006 / viro.1998.9028.
CAS PubMed Google Scholar
Ди Марцио П., Чоу С., Эбрайт М., Кноблаух Р., Ландау Н.Р.: Мутационный анализ остановки клеточного цикла, ядерной локализации и упаковки вирионов вируса иммунодефицита человека типа 1 Vpr. J Virol. 1995, 69: 7909-7916.
PubMed Central CAS PubMed Google Scholar
Jenkins Y, McEntee M, Weis K, Greene WC: Характеристика ядерного импорта vpr ВИЧ-1: анализ сигналов и путей. J Cell Biol. 1998, 143: 875-885.10.1083 / jcb.143.4.875.
PubMed Central CAS PubMed Google Scholar
Subbramanian RA, Yao XJ, Dilhuydy H, Rougeau N, Bergeron D, Robitaille Y, Cohen EA: Локализация Vpr вируса иммунодефицита человека типа 1: ядерный транспорт вирусного белка, модулируемый предполагаемой амфипатической спиральной структурой и ее отношение к биологической активности. J Mol Biol. 1998, 278: 13-30. 10.1006 / jmbi.1998.1685.
CAS PubMed Google Scholar
Карни О., Фридлер А., Закай Н., Гилон С., Лойтер А: пептид, полученный из N-концевой области Vpr ВИЧ-1, способствует ядерному импорту в проницаемые клетки: выяснение области NLS Vpr. FEBS Lett. 1998, 429: 421-425. 10.1016 / S0014-5793 (98) 00645-0.
CAS PubMed Google Scholar
Букринский М.И., Хаффар ОК: ядерный импорт ВИЧ-1: матричный белок снова в центре внимания, на этот раз вместе с Vpr. Mol Med.1998, 4: 138-143.
PubMed Central CAS PubMed Google Scholar
Агостини И., Попов С., Ли Дж., Дубровский Л., Хао Т., Букринский М.: Белок теплового шока 70 может заменить вирусный белок R ВИЧ-1 во время ядерного импорта вирусного прединтеграционного комплекса. Exp Cell Res. 2000, 259: 398-403. 10.1006 / excr.2000.4992.
CAS PubMed Google Scholar
Zennou V, Petit C, Guetard D, Nerhbass U, Montagnier L, Charneau P: ядерный импорт генома ВИЧ-1 опосредуется центральным лоскутом ДНК. Клетка. 2000, 101: 173-185. 10.1016 / S0092-8674 (00) 80828-4.
CAS PubMed Google Scholar
Шерман М.П., Грин WC: Проскользнув в дверь: проникновение ВИЧ в ядро. Микробы заражают. 2002, 4: 67-73. 10.1016 / S1286-4579 (01) 01511-8.
CAS PubMed Google Scholar
Шерман М.П., де Норонья С.М., Хеуш М.И., Грин С., Грин В.С.: Нуклеоцитоплазматическое перемещение вируса иммунодефицита человека типа 1 Vpr. J Virol. 2001, 75: 1522-1532. 10.1128 / JVI.75.3.1522-1532.2001.
PubMed Central CAS PubMed Google Scholar
Sherman MP, de Noronha CM, Eckstein LA, Hataye J, Mundt P, Williams SA, Neidleman JA, Goldsmith MA, Greene WC: Ядерный экспорт Vpr необходим для эффективной репликации вируса иммунодефицита человека типа 1 в тканевые макрофаги.J Virol. 2003, 77: 7582-7589. 10.1128 / JVI.77.13.7582-7589.2003.
PubMed Central CAS PubMed Google Scholar
Jowett JB, Planelles V, Poon B, Shah NP, Chen ML, Chen IS: Ген vpr вируса иммунодефицита человека типа 1 задерживает инфицированные Т-клетки в фазе G2 + M клеточного цикла. J Virol. 1995, 69: 6304-6313.
PubMed Central CAS PubMed Google Scholar
He J, Choe S, Walker R, Di Marzio P, Morgan DO, Landau NR: вирусный белок R (Vpr) вируса иммунодефицита человека типа 1 задерживает клетки в фазе G2 клеточного цикла, ингибируя активность p34cdc2. J Virol. 1995, 69: 6705-6711.
PubMed Central CAS PubMed Google Scholar
Re F, Braaten D, Franke EK, Luban J: Vpr вируса иммунодефицита человека типа 1 останавливает клеточный цикл в G2, ингибируя активацию p34cdc2-cyclin B.J Virol. 1995, 69: 6859-6864.
PubMed Central CAS PubMed Google Scholar
Bartz SR, Rogel ME, Emerman M: Контроль клеточного цикла вируса иммунодефицита человека типа 1: Vpr является цитостатическим и опосредует накопление G2 по механизму, который отличается от контроля контрольных точек повреждения ДНК. J Virol. 1996, 70: 2324-2331.
PubMed Central CAS PubMed Google Scholar
Planelles V, Jowett JB, Li QX, Xie Y, Hahn B, Chen IS: Vpr-индуцированная остановка клеточного цикла сохраняется среди лентивирусов приматов. J Virol. 1996, 70: 2516-2524.
PubMed Central CAS PubMed Google Scholar
Пун Б., Гровит-Фербас К., Стюарт С.А., Чен ISY: остановка клеточного цикла с помощью Vpr в вирионах ВИЧ-1 и нечувствительность к антиретровирусным агентам. Наука. 1998, 281: 266-269. 10.1126 / science.281.5374.266.
CAS PubMed Google Scholar
Чжан Ц., Расмуссен Ц., Чанг Л.Дж .: Эффекты ингибирования клеточного цикла ВИЧ и SIV Vpr и Vpx в дрожжах Schizosaccharomyces pombe. Вирусология. 1997, 230: 103-112. 10.1006 / viro.1997.8459.
CAS PubMed Google Scholar
Zhao Y, Cao J, O’Gorman MR, Yu M, Yogev R: Влияние экспрессии гена белка R (vpr) вируса иммунодефицита человека типа 1 на основную клеточную функцию делящихся дрожжей Schizosaccharomyces pombe. J Virol.1996, 70: 5821-5826.
PubMed Central CAS PubMed Google Scholar
Яо XJ, Lemay J, Rougeau N, Clement M, Kurtz S, Belhumeur P, Cohen EA: Генетический отбор пептидных ингибиторов вируса иммунодефицита человека типа 1 Vpr. J Biol Chem. 2002, 277: 48816-48826. 10.1074 / jbc.M207982200.
CAS PubMed Google Scholar
Bouzar AB, Guiguen F, Morin T, Villet S, Fornazero C, Garnier C, Gallay K, Gounel F, Favier C, Durand J и др.: Специфическая задержка G2 козьих клеток, инфицированных козьим артритом вирус энцефалита, экспрессирующий гены vpr и vpx вируса иммунодефицита обезьян.Вирусология. 2003, 309: 41-52. 10.1016 / S0042-6822 (03) 00014-Х.
CAS PubMed Google Scholar
Goh WC, Rogel ME, Kinsey CM, Michael SF, Fultz PN, Nowak MA, Hahn BH, Emerman M: ВИЧ-1 Vpr увеличивает вирусную экспрессию путем манипулирования клеточным циклом: механизм отбора Vpr in vivo. Nat Med. 1998, 4: 65-71. 10.1038 / нм0198-065.
CAS PubMed Google Scholar
Чжоу И, Ратнер Л.: Фосфорилирование вируса иммунодефицита человека типа 1 Vpr регулирует остановку клеточного цикла. J Virol. 2000, 74: 6520-6527. 10.1128 / JVI.74.14.6520-6527.2000.
PubMed Central CAS PubMed Google Scholar
Агостини И., Попов С., Хао Т., Ли Дж. Х., Дубровский Л., Чайка О., Чайка Н., Льюис Р., Букринский М.: Фосфорилирование Vpr регулирует ядерный импорт 1 типа ВИЧ и макрофагальную инфекцию. AIDS Res Hum Retroviruses.2002, 18: 283-288. 10.1089 / 088
2753472856.CAS PubMed Google Scholar
Медсестра П: КПП достигли совершеннолетия. Клетка. 1997, 91: 865-867. 10.1016 / S0092-8674 (00) 80476-6.
CAS PubMed Google Scholar
Смитс В.А., Медема Р.Х .: Проверка перехода G (2) / M. Biochim Biophys Acta. 2001, 1519: 1-12.
CAS PubMed Google Scholar
Mahalingam S, Ayyavoo V, Patel M, Kieber-Emmons T, Kao GD, Muschel RJ, Weiner DB: Vpr ВИЧ-1 взаимодействует с человеческим гомологом mov34 массой 34 кДа, клеточным фактором, связанным с фазовым переходом G2 / M клеточный цикл млекопитающих. Proc Natl Acad Sci U S. A. 1998, 95: 3419-3424. 10.1073 / pnas.95.7.3419.
PubMed Central CAS PubMed Google Scholar
Ramanathan MP, Curley E, Su M, Chambers JA, Weiner DB: Карбоксильный конец hVIP / mov34 имеет решающее значение для взаимодействия ВИЧ-1-Vpr и передачи сигналов, опосредованной глюкокортикоидами.J Biol Chem. 2002, 277: 47854-47860. 10.1074 / jbc.M2030.
CAS PubMed Google Scholar
Грагеров А., Кино Т., Ильина-Грагерова Г., Хрусос Г. П., Павлакис Г. Н.: HHR23A, человеческий гомолог дрожжевого репаративного белка RAD23, специфически взаимодействует с белком Vpr и предотвращает остановку клеточного цикла, но не транскрипционные эффекты Впр. Вирусология. 1998, 245: 323-330. 10.1006 / viro.1998.9138.
CAS PubMed Google Scholar
Withers-Ward ES, Jowett JB, Stewart SA, Xie YM, Garfinkel A, Shibagaki Y, Chow SA, Shah N, Hanaoka F, Sawitz DG и др .: Vpr вируса иммунодефицита человека типа 1 взаимодействует с HHR23A, вовлеченным клеточным белком в репарации ДНК с эксцизией нуклеотидов. J Virol. 1997, 71: 9732-9742.
PubMed Central CAS PubMed Google Scholar
Bouhamdan M, Benichou S, Rey F, Navarro JM, Agostini I, Spire B, Camonis J, Slupphaug G, Vigne R, Benarous R, Sire J: белок Vpr вируса иммунодефицита человека 1 типа связывается с урацилом ДНК-гликозилаза, фермент репарации ДНК.J Virol. 1996, 70: 697-704.
PubMed Central CAS PubMed Google Scholar
Mansky LM, Preveral S, Le Rouzic E, Bernard LC, Selig L, Depienne C, Benarous R, Benichou S: Взаимодействие Vpr вируса иммунодефицита человека типа 1 с репарационным белком HHR23A не коррелирует с множественными биологические функции Впр. Вирусология. 2001, 282: 176-185. 10.1006 / viro.2000.0791.
CAS PubMed Google Scholar
Selig L, Benichou S, Rogel ME, Wu LI, Vodicka MA, Sire J, Benarous R, Emerman M: ДНК-гликозилаза урацила специфически взаимодействует с Vpr как вируса иммунодефицита человека типа 1, так и вируса иммунодефицита обезьян, покрытых сажей, но связывание действует не коррелируют с остановкой клеточного цикла. J Virol. 1997, 71: 4842-4846.
PubMed Central CAS PubMed Google Scholar
Пун Б., Джоветт Дж. Б., Стюарт С.А., Армстронг Р.В., Риштон Г.М., Чен И.С.: Ген vpr вируса иммунодефицита человека типа 1 вызывает фенотипические эффекты, аналогичные эффектам алкилирующего агента ДНК, азотистого иприта.J Virol. 1997, 71: 3961-3971.
PubMed Central CAS PubMed Google Scholar
Cliby WA, Lewis KA, Lilly KK, Kaufmann SH: S-фаза и задержка G2, вызванная ядами топоизомеразы I, зависит от функции киназы ATR. J Biol Chem. 2002, 277: 1599-1606. 10.1074 / jbc.M106287200.
CAS PubMed Google Scholar
Roshal M, Kim B, Zhu Y, Nghiem P, Planelles V: Активация ATR-опосредованного ответа на повреждение ДНК вирусным белком ВИЧ-1 R.J Biol Chem. 2003, 278: 25879-25886. 10.1074 / jbc.M303948200.
CAS PubMed Google Scholar
Hrimech M, Yao XJ, Branton PE, Cohen EA: Vpr-опосредованная остановка клеточного цикла G (2) вируса иммунодефицита человека типа 1: Vpr вмешивается в сигнальные каскады клеточного цикла, взаимодействуя с B-субъединицей серина / треониновая протеинфосфатаза 2А. Эмбо Дж. 2000, 19: 3956-3967. 10.1093 / emboj / 19.15.3956.
PubMed Central CAS PubMed Google Scholar
Zolnierowicz S: протеинфосфатаза типа 2A, комплексный регулятор многочисленных сигнальных путей. Biochem Pharmacol. 2000, 60: 1225-1235. 10.1016 / S0006-2952 (00) 00424-Х.
CAS PubMed Google Scholar
Масуда М., Нагаи Ю., Осима Н., Танака К., Мураками Х., Игараши Х., Окаяма Х .: Генетические исследования с делящимися дрожжами Schizosaccharomyces pombe предполагают участие wee1, ppa2 и rad24 в индукции остановки клеточного цикла. вирусом иммунодефицита человека 1 типа Vpr.J Virol. 2000, 74: 2636-2646. 10.1128 / JVI.74.6.2636-2646.2000.
PubMed Central CAS PubMed Google Scholar
Хагтинг А., Джекман М., Симпсон К., Пайнс Дж. Для транслокации циклина В1 в ядро в профазе требуется сигнал импорта ядра, зависимый от фосфорилирования. Curr Biol. 1999, 9: 680-689. 10.1016 / S0960-9822 (99) 80308-Х.
CAS PubMed Google Scholar
Карр AM: Молекулярная биология. Начиная с конца. Наука. 2003, 300: 1512-1513. 10.1126 / science.1085689.
CAS PubMed Google Scholar
Hutchison CJ: Ламины: строительные блоки или регуляторы экспрессии генов ?. Nat Rev Mol Cell Biol. 2002, 3: 848-858. 10.1038 / nrm950.
CAS PubMed Google Scholar
Алимонти Дж. Б., Болл ТБ, Фоук К.Р.: Механизмы гибели CD4 + Т-лимфоцитов при инфицировании вирусом иммунодефицита человека и СПИДе.J Gen Virol. 2003, 84: 1649-1661. 10.1099 / vir.0.19110-0.
CAS PubMed Google Scholar
Ясуда Дж., Мияо Т., Камата М., Аида Ю., Ивакура Ю. Апоптоз Т-клеток вызывает истощение периферических Т-клеток у мышей, трансгенных по гену vpr ВИЧ-1. Вирусология. 2001, 285: 181-192. 10.1006 / viro.2001.0964.
CAS PubMed Google Scholar
Ayyavoo V, Mahboubi A, Mahalingam S, Ramalingam R, Kudchodkar S, Williams WV, Green DR, Weiner DB: Vpr ВИЧ-1 подавляет активацию иммунной системы и апоптоз посредством регуляции ядерного фактора каппа B [см. Комментарии] .Nat Med. 1997, 3: 1117-1123. 10,1038 / нм1097-1117.
CAS PubMed Google Scholar
Conti L, Rainaldi G, Matarrese P, Varano B, Rivabene R, Columba S, Sato A, Belardelli F, Malorni W., Gessani S: белок vpr ВИЧ-1 действует как негативный регулятор апоптоза в линия лимфобластоидных Т-клеток человека: возможные последствия для патогенеза СПИДа. J Exp Med. 1998, 187: 403-413. 10.1084 / jem.187.3.403.
PubMed Central CAS PubMed Google Scholar
Стюарт С.А., Пун Б., Джоветт Дж. Б., Чен И.С.: Vpr вируса иммунодефицита человека типа 1 индуцирует апоптоз после остановки клеточного цикла. J Virol. 1997, 71: 5579-5592.
PubMed Central CAS PubMed Google Scholar
Стюарт С.А., Пун Б., Сонг Дж.Й., Чен И.С.: vpr вируса иммунодефицита человека типа 1 индуцирует апоптоз через активацию каспазы. J Virol. 2000, 74: 3105-3111. 10.1128 / JVI.74.7.3105-3111.2000.
PubMed Central CAS PubMed Google Scholar
Watanabe N, Yamaguchi T., Akimoto Y, Rattner JB, Hirano H, Nakauchi H: индукция остановки M-фазы и апоптоза после экспрессии Vpr ВИЧ-1 посредством разобщения ядерного и центросомного цикла в клетках HeLa. Exp Cell Res. 2000, 258: 261-269. 10.1006 / excr.2000.4908.
CAS PubMed Google Scholar
Yao XJ, Mouland AJ, Subbramanian RA, Forget J, Rougeau N, Bergeron D, Cohen EA: Vpr стимулирует вирусную экспрессию и индуцирует гибель клеток в делящихся Т-клетках Jurkat, инфицированных вирусом иммунодефицита человека 1-го типа.J Virol. 1998, 72: 4686-4693.
PubMed Central CAS PubMed Google Scholar
Muthumani K, Hwang DS, Desai BM, Zhang D, Dayes N, Green DR, Weiner DB: Vpr ВИЧ-1 индуцирует апоптоз через каспазу 9 в Т-клетках и мононуклеарных клетках периферической крови. J Biol Chem. 2002, 277: 37820-37831. 10.1074 / jbc.M205313200.
CAS PubMed Google Scholar
Nishizawa M, Kamata M, Katsumata R, Aida Y: усеченная на карбокси-конце форма белка Vpr вируса иммунодефицита человека типа 1 индуцирует апоптоз посредством остановки клеточного цикла G (1). J Virol. 2000, 74: 6058-6067. 10.1128 / JVI.74.13.6058-6067.2000.
PubMed Central CAS PubMed Google Scholar
Nishizawa M, Kamata M, Mojin T., Nakai Y, Aida Y: индукция апоптоза белком Vpr вируса иммунодефицита человека типа 1 происходит независимо от G (2) остановки клеточного цикла.Вирусология. 2000, 276: 16-26. 10.1006 / viro.2000.0534.
CAS PubMed Google Scholar
Юань Х., Се Ю.М., Чен И.С.: Истощение киназы Wee-1 необходимо как для Vpr- вируса иммунодефицита человека типа 1, так и для апоптоза, индуцированного гамма-облучением. J Virol. 2003, 77: 2063-2070. 10.1128 / JVI.77.3.2063-2070.2003.
PubMed Central CAS PubMed Google Scholar
Крек В., Нигг Е.А.: Мутации сайтов фосфорилирования p34cdc2 вызывают преждевременные митотические события в клетках HeLa: доказательства двойного блока активации киназы p34cdc2 у позвоночных. Embo J. 1991, 10: 3331-3341.
PubMed Central CAS PubMed Google Scholar
Heald R, McLoughlin M, McKeon F: Human wee1 поддерживает митотическое время, защищая ядро от цитоплазматически активированной киназы Cdc2. Клетка. 1993, 74: 463-474.10.1016 / 0092-8674 (93) 80048-Дж.
CAS PubMed Google Scholar
Чанг Ф., Ре Ф., Себастьян С., Сазер С., Любан Дж. Vpr ВИЧ-1 вызывает дефекты митоза, цитокинеза, ядерной структуры и центросом. Mol Biol Cell. 2004, 15: 1793-1801. 10.1091 / mbc.E03-09-0691.
PubMed Central CAS PubMed Google Scholar
Jacotot E, Ferri KF, El Hamel C, Brenner C, Druillennec S, Hoebeke J, Rustin P, Metivier D, Lenoir C, Geuskens M, et al: Контроль проницаемости митохондриальной мембраны с помощью взаимодействия транслокатора аденинового нуклеотида с вирусным белком ВИЧ-1 rR и Bcl-2.J Exp Med. 2001, 193: 509-519. 10.1084 / jem.193.4.509.
PubMed Central CAS PubMed Google Scholar
Ван X: Расширяющаяся роль митохондрий в апоптозе. Genes Dev. 2001, 15: 2922-2933.
CAS PubMed Google Scholar
Roumier T, Vieira HL, Castedo M, Ferri KF, Boya P, Andreau K, Druillennec S, Joza N, Penninger JM, Roques B, Kroemer G: С-концевой фрагмент Vpr ВИЧ-1 индуцирует гибель клеток через каспазонезависимый митохондриальный путь.Смерть клетки отличается. 2002, 9: 1212-1219. 10.1038 / sj.cdd.4401089.
CAS PubMed Google Scholar
Muthumani K, Choo AY, Hwang DS, Chattergoon MA, Dayes NN, Zhang D, Lee MD, Duvvuri U, Weiner DB: Механизм апоптоза, индуцированного вирусным белком ВИЧ-1 R. Biochem Biophys Res Commun. 2003, 304: 583-592. 10.1016 / S0006-291X (03) 00631-4.
CAS PubMed Google Scholar
Рошаль М., Чжу Ю., Planelles V: Апоптоз при СПИДе. Апоптоз. 2001, 6: 103-116. 10.1023 / А: 1009636530839.
CAS PubMed Google Scholar
Lum JJ, Cohen OJ, Nie Z, Weaver JG, Gomez TS, Yao XJ, Lynch D, Pilon AA, Hawley N, Kim JE и др.: Vpr R77Q связан с длительно непрогрессирующей ВИЧ-инфекцией. и нарушение индукции апоптоза. J Clin Invest. 2003, 111: 1547-1554. 10.1172 / JCI200316233.
PubMed Central CAS PubMed Google Scholar
Jian H, Zhao LJ: Проапоптотическая активность вспомогательного регуляторного белка ВИЧ-1 Vpr зависит от подтипа и сильно усиливается неконсервативными изменениями остатка лейцина в положении 64. J Biol Chem. 2003, 278: 44326-44330. 10.1074 / jbc.C300378200.
CAS PubMed Google Scholar
Леви Д.Н., Рафаэли Ю., Вайнер ДБ: Внеклеточный белок Vpr увеличивает клеточную проницаемость для репликации вируса иммунодефицита человека и реактивирует вирус с латентного периода.J Virol. 1995, 69: 1243-1252.
PubMed Central CAS PubMed Google Scholar
Levy DN, Refaeli Y, MacGregor RR, Weiner DB: сывороточный Vpr регулирует продуктивную инфекцию и латентный период вируса иммунодефицита человека типа 1. Proc Natl Acad Sci U S. A. 1994, 91: 10873-10877.
PubMed Central CAS PubMed Google Scholar
Патель С.А., Мухтар М., Харли С., Кулькоски Дж., Померанц Р.Дж .: Лентивирусная экспрессия Vpr ВИЧ-1 вызывает апоптоз в нейронах человека.J Neurovirol. 2002, 8: 86-99. 10.1080 / 135502802552.
CAS PubMed Google Scholar
Piller SC, Ewart GD, Jans DA, Gage PW, Cox GB: Аминоконцевая область Vpr вируса иммунодефицита человека типа 1 образует ионные каналы и убивает нейроны. J Virol. 1999, 73: 4230-4238.
PubMed Central CAS PubMed Google Scholar
Huang MB, Weeks O, Zhao LJ, Saltarelli M, Bond VC: эффекты внеклеточного белка vpr вируса иммунодефицита человека типа 1 в первичных культурах кортикальных клеток крыс.J Neurovirol. 2000, 6: 202-220.
CAS PubMed Google Scholar
Piller SC, Ewart GD, Premkumar A, Cox GB, Gage PW: белок Vpr вируса иммунодефицита человека типа 1 образует катион-селективные каналы в плоских липидных бислоях. Proc Natl Acad Sci U S. A. 1996, 93: 111-115. 10.1073 / pnas.93.1.111.
PubMed Central CAS PubMed Google Scholar
Патель К.А., Мухтар М., Померанц Р.Дж.: Vpr вируса иммунодефицита человека типа 1 индуцирует апоптоз в нейрональных клетках человека. J Virol. 2000, 74: 9717-9726. 10.1128 / JVI.74.20.9717-9726.2000.
PubMed Central CAS PubMed Google Scholar
Cohen EA, Dehni G, Sodroski JG, Haseltine WA: продукт vpr вируса иммунодефицита человека представляет собой связанный с вирионом регуляторный белок. J Virol. 1990, 64: 3097-3099.
PubMed Central CAS PubMed Google Scholar
Огава К., Сибата Р., Киёмасу Т., Хигучи И., Кишида Ю., Ишимото А., Адачи А.: Мутационный анализ открытой рамки считывания vpr вируса иммунодефицита человека. J Virol. 1989, 63: 4110-4114.
PubMed Central CAS PubMed Google Scholar
Felzien LK, Woffendin C, Hottiger MO, Subbramanian RA, Cohen EA, Nabel GJ: Активация транскрипции ВИЧ дополнительным белком, VPR, опосредуется коактиватором p300.Proc Natl Acad Sci U S. A. 1998, 95: 5281-5286. 10.1073 / pnas.95.9.5281.
PubMed Central CAS PubMed Google Scholar
Gummuluru S, Emerman M: Опосредованная клеточным циклом и Vpr регуляция экспрессии вируса иммунодефицита человека типа 1 в первичных и трансформированных линиях Т-клеток. J Virol. 1999, 73: 5422-5430.
PubMed Central CAS PubMed Google Scholar
Subbramanian RA, Kessous-Elbaz A, Lodge R, Forget J, Yao XJ, Bergeron D, Cohen EA: Vpr вируса иммунодефицита человека типа 1 является положительным регулятором вирусной транскрипции и инфекционности в первичных макрофагах человека. J Exp Med. 1998, 187: 1103-1111. 10.1084 / jem.187.7.1103.
PubMed Central CAS PubMed Google Scholar
Hrimech M, Yao XJ, Bachand F, Rougeau N, Cohen EA: Vpr вируса иммунодефицита человека типа 1 (ВИЧ-1) функционирует как ранний белок во время ВИЧ-1 инфекции.J Virol. 1999, 73: 4101-4109.
PubMed Central CAS PubMed Google Scholar
Hogan TH, Nonnemacher MR, Krebs FC, Henderson A, Wigdahl B: Связывание Vpr ВИЧ-1 с цис-действующими элементами LTR C / EBP ВИЧ-1 и прилегающими областями является последовательноспецифичным. Biomed Pharmacother. 2003, 57: 41-48. 10.1016 / S0753-3322 (02) 00333-5.
CAS PubMed Google Scholar
Ван Л., Мукерджи С., Цзя Ф., Нараян О., Чжао Л.Дж .: Взаимодействие вирионного белка Vpr вируса иммунодефицита человека типа 1 с клеточным транскрипционным фактором Sp1 и трансактивация длинного концевого повтора вируса. J Biol Chem. 1995, 270: 25564-25569. 10.1074 / jbc.270.43.25564.
CAS PubMed Google Scholar
Vanitharani R, Mahalingam S, Rafaeli Y, Singh SP, Srinivasan A, Weiner DB, Ayyavoo V: Vpr ВИЧ-1 трансактивирует LTR-направленную экспрессию через последовательности, присутствующие в пределах от -278 до -176, и увеличивает репликацию вируса в пробирка.Вирусология. 2001, 289: 334-342. 10.1006 / viro.2001.1153.
CAS PubMed Google Scholar
Пун Б, Чен И.С.: Vpr вируса иммунодефицита человека типа 1 (ВИЧ-1) усиливает экспрессию неинтегрированной ДНК ВИЧ-1. J Virol. 2003, 77: 3962-3972. 10.1128 / JVI.77.7.3962-3972.2003.
PubMed Central CAS PubMed Google Scholar
Zhu Y, Roshal M, Li F, Blackett J, Planelles V: Повышение регуляции сурвивина с помощью HIV-1 Vpr.Апоптоз. 2003, 8: 71-79. 10.1023 / А: 1021653119934.
CAS PubMed Google Scholar
Roux P, Alfieri C, Hrimech M, Cohen EA, Tanner JE: Активация факторов транскрипции NF-kappaB и NF-IL-6 белком R вируса иммунодефицита человека 1 типа (Vpr) индуцирует экспрессию интерлейкина-8. . J Virol. 2000, 74: 4658-4665. 10.1128 / JVI.74.10.4658-4665.2000.
PubMed Central CAS PubMed Google Scholar
Chowdhury IH, Wang XF, Landau NR, Robb ML, Polonis VR, Birx DL, Kim JH: ВИЧ-1 Vpr активирует ингибитор клеточного цикла p21 / Waf1 / Cip1: потенциальный механизм остановки клеточного цикла G2 / M. Вирусология. 2003, 305: 371-377. 10.1006 / viro.2002.1777.
CAS PubMed Google Scholar
Кино Т., Грагеров А., Копп Дж. Б., Штаубер Р. Х., Павлакис Г. Н., Хрусос Г. П.: Вирион-ассоциированный белок vpr ВИЧ-1 является коактиватором рецептора глюкокортикоидов человека.J Exp Med. 1999, 189: 51-62. 10.1084 / jem.189.1.51.
PubMed Central CAS PubMed Google Scholar
Рафаэли Ю., Леви Д. Н., Вейнер Д. Б.: Комплекс глюкокортикоидного рецептора типа II является мишенью продукта гена vpr ВИЧ-1. Proc Natl Acad Sci U S. A. 1995, 92: 3621-3625.
PubMed Central CAS PubMed Google Scholar
Hottiger MO, Nabel GJ: Репликация вирусов и коактиваторы p300 и CBP.Trends Microbiol. 2000, 8: 560-565. 10.1016 / S0966-842X (00) 01874-6.
CAS PubMed Google Scholar
Agostini I, Navarro JM, Rey F, Bouhamdan M, Spire B, Vigne R, Sire J: Трансактиватор Vpr вируса иммунодефицита человека типа 1: взаимодействие с промоторными доменами активатора и связывание с TFIIB. J Mol Biol. 1996, 261: 599-606. 10.1006 / jmbi.1996.0485.
CAS PubMed Google Scholar
Савая Б.Е., Халили К., Мерсер В.Е., Денисова Л., Амини С.: Совместные действия ВИЧ-1 Vpr и p53 модулируют транскрипцию вирусных генов. J Biol Chem. 1998, 273: 20052-20057. 10.1074 / jbc.273.32.20052.
CAS PubMed Google Scholar
Кино Т., Грагеров А., Слободская О., Цопаномичалоу М., Хрусос Г. П., Павлакис Г. Н.: Дополнительный белок вируса иммунодефицита человека типа 1 (ВИЧ-1) Vpr индуцирует транскрипцию промоторов, реагирующих на ВИЧ-1 и глюкокортикоиды, посредством связывание непосредственно с коактиваторами p300 / CBP.J Virol. 2002, 76: 9724-9734. 10.1128 / JVI.76.19.9724-9734.2002.
PubMed Central CAS PubMed Google Scholar
Шерман М.П., де Норонья К.М., Пирс Д., Грин В.К.: vpr вируса иммунодефицита человека типа 1 содержит две богатые лейцином спирали, которые опосредуют коактивацию рецептора глюкокортикоидов независимо от его эффектов на остановку клеточного цикла G (2) [In Цитирование процесса. J Virol. 2000, 74: 8159-8165. 10.1128 / JVI.74.17.8159-8165.2000.
PubMed Central CAS PubMed Google Scholar
Janket ML, Manickam P, Majumder B, Thotala D, Wagner M, Schafer EA, Collman RG, Srinivasan A, Ayyavoo V: Дифференциальная регуляция клеточных генов хозяина вирусным белком ВИЧ-1 R (Vpr): Анализ микрочипов кДНК с использованием изогенного вируса.